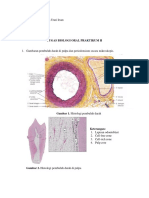
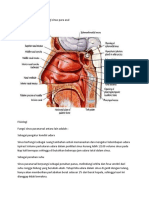
Anda mungkin juga menyukai
- Dasar TransportasiDokumen14 halamanDasar TransportasiAbraham BoltalBelum ada peringkat
- MIKROSIRKULASIDokumen6 halamanMIKROSIRKULASILempahBelatjanBelum ada peringkat
- EndoteliumDokumen28 halamanEndoteliumDesty100% (2)
- TUGAS Sel EndotheliumDokumen12 halamanTUGAS Sel Endotheliumcandra agusdiBelum ada peringkat
- Pertukaran Cairan Mikrovaskuler Dan Terapi CairanDokumen28 halamanPertukaran Cairan Mikrovaskuler Dan Terapi CairanWanAzrenBelum ada peringkat
- Laporan Mikrosirkulasi JadiDokumen20 halamanLaporan Mikrosirkulasi JadiDwi Ayu NingtyasBelum ada peringkat
- Kelompok 1Dokumen58 halamanKelompok 1nikkennnBelum ada peringkat
- Pertukaran Cairan MikrovaskularDokumen7 halamanPertukaran Cairan MikrovaskularodzieBelum ada peringkat
- Re Dan GolgiDokumen30 halamanRe Dan Golgishofa_adityaBelum ada peringkat
- Ringksan Individu BSM1Dokumen2 halamanRingksan Individu BSM1Digdo SudigyoBelum ada peringkat
- Rekap SPODokumen169 halamanRekap SPOShinta Dewi SiregarBelum ada peringkat
- PAPER BIOMEKANIKA Dan BIOTRANSPORTASIDokumen20 halamanPAPER BIOMEKANIKA Dan BIOTRANSPORTASIFauziyah Firdausi M. S0% (1)
- Makalah Transpor SelDokumen8 halamanMakalah Transpor SelMuhammad AminBelum ada peringkat
- Blood Ocular BarrierDokumen53 halamanBlood Ocular Barrieraffan azhari100% (1)
- Endometrium Dan MiometriumDokumen112 halamanEndometrium Dan MiometriumNurmailis SelayanBelum ada peringkat
- Membrane TransportDokumen13 halamanMembrane TransportSitti Rahmah MustakinBelum ada peringkat
- Bedah Buku Flap NewDokumen13 halamanBedah Buku Flap NewmiadexxBelum ada peringkat
- Brain Blood Barrier (Sawar Darah Otak) : PendahuluanDokumen17 halamanBrain Blood Barrier (Sawar Darah Otak) : PendahuluanbobirfansyahputraBelum ada peringkat
- Mekanisme Pengantaran Obat Ke OtakDokumen3 halamanMekanisme Pengantaran Obat Ke OtakRoza EfdanikaBelum ada peringkat
- BiotransperDokumen8 halamanBiotransperdyah prashantiBelum ada peringkat
- Makalah Biologi KLPK 3Dokumen14 halamanMakalah Biologi KLPK 3Makmur S. SosBelum ada peringkat
- Bab 11dkDokumen143 halamanBab 11dkmarthagusnadiBelum ada peringkat
- Wrap Up EdemaDokumen25 halamanWrap Up Edemanandakusyuda100% (3)
- A - 009 - Risyanuari Saufika Utari Irsan - MikrosirkulasiDokumen5 halamanA - 009 - Risyanuari Saufika Utari Irsan - Mikrosirkulasirisyanuari saufikaBelum ada peringkat
- Patologi 4ADokumen20 halamanPatologi 4Arahmant9162Belum ada peringkat
- Anatomi P. Darah Dan HistologiDokumen16 halamanAnatomi P. Darah Dan HistologiAdelBelum ada peringkat
- Pemilihan Cairan Resusitasi Untuk Pertahankan Endotelial GlycocalyxDokumen16 halamanPemilihan Cairan Resusitasi Untuk Pertahankan Endotelial GlycocalyxtwlrBelum ada peringkat
- 20031086-Nadiyah Farhah Salsabila-Laporan Praktikum Permeabilitas SelDokumen10 halaman20031086-Nadiyah Farhah Salsabila-Laporan Praktikum Permeabilitas SelNadiyah SalsabilahBelum ada peringkat
- Kontraksi UterusDokumen11 halamanKontraksi UterusMuhammad Fadhil100% (1)
- MAKALAH Transportasi ZatDokumen14 halamanMAKALAH Transportasi ZatAchmad A'fhiecc100% (2)
- 4B1 - Kelompok 3 - Resume Jurnal Jaringan Parenkim PDFDokumen4 halaman4B1 - Kelompok 3 - Resume Jurnal Jaringan Parenkim PDFRAHIMA SALSABILA 2021Belum ada peringkat
- Tugas Fisiologi Ppds Bedah Maret 2020Dokumen75 halamanTugas Fisiologi Ppds Bedah Maret 2020Ibnu CaesarBelum ada peringkat
- Makalah Membran Sel (Un-Edited)Dokumen7 halamanMakalah Membran Sel (Un-Edited)Rizki ChandraBelum ada peringkat
- Mekanisme Transpor Membran SelDokumen3 halamanMekanisme Transpor Membran Seleko widodoBelum ada peringkat
- Mekanisme Transpor Pada Membran PlasmaDokumen9 halamanMekanisme Transpor Pada Membran PlasmaFionikaBelum ada peringkat
- Tugas KLP Genap Prof Arni Biologi Sel (Membran Sel)Dokumen6 halamanTugas KLP Genap Prof Arni Biologi Sel (Membran Sel)nadya khaira nurdiBelum ada peringkat
- Mikrosirkulasi Dan LimfatikDokumen11 halamanMikrosirkulasi Dan LimfatikMuhammad AsyrafBelum ada peringkat
- Tugas Fartok I Kelompok 14Dokumen8 halamanTugas Fartok I Kelompok 14Alif SyahabBelum ada peringkat
- Pertukaran Zat Antara Sle Dengan Lingkungannya Terjadi Karena Adanya Mekanisme Transpor Zat Melalui Membran Sel Yang Berlangsung Secara DifusiDokumen3 halamanPertukaran Zat Antara Sle Dengan Lingkungannya Terjadi Karena Adanya Mekanisme Transpor Zat Melalui Membran Sel Yang Berlangsung Secara DifusiRenita IndrianiBelum ada peringkat
- Kesimpulan Exchange MikrovaskularDokumen8 halamanKesimpulan Exchange MikrovaskularRifqi FuadiBelum ada peringkat
- Fisiologi GinjalDokumen6 halamanFisiologi GinjalRaisaBelum ada peringkat
- Laporan MikroDokumen21 halamanLaporan MikroAndi Sitti Hajrah YusufBelum ada peringkat
- DWIPRAYOGO WIBOWO - KARBOHIDRAT TUGAS KELOMPOK-with-cover-page-v2Dokumen13 halamanDWIPRAYOGO WIBOWO - KARBOHIDRAT TUGAS KELOMPOK-with-cover-page-v2Yizlia FebiBelum ada peringkat
- Fisiologi Sel, Homeostasis Dan Interaksi Sel-BCBBiomedik-PW2013Dokumen81 halamanFisiologi Sel, Homeostasis Dan Interaksi Sel-BCBBiomedik-PW2013Surajul AudaBelum ada peringkat
- Histologi Esofagus Dan GasterDokumen7 halamanHistologi Esofagus Dan GasterDelvi AkechiBelum ada peringkat
- GlomerulonefritisDokumen29 halamanGlomerulonefritisnitapggBelum ada peringkat
- Sitohisto 1Dokumen6 halamanSitohisto 1NindyawatiBelum ada peringkat
- Wrap Up Skenario 2 BengkakDokumen17 halamanWrap Up Skenario 2 BengkaksintamirosmalindaBelum ada peringkat
- Struktur Sel Tumbuhan Dan HewanDokumen6 halamanStruktur Sel Tumbuhan Dan HewanMeyLaa KrisnaBelum ada peringkat
- Ion Kalsium Pada Metabolisme Matriks EkstrselulerDokumen37 halamanIon Kalsium Pada Metabolisme Matriks EkstrselulerKris RatuBelum ada peringkat
- Makalah Sel KLP 3Dokumen19 halamanMakalah Sel KLP 3Robi AshadBelum ada peringkat
- Makalah Biologi SelDokumen15 halamanMakalah Biologi SelBendahara BarangBelum ada peringkat
- Makalah Tentang Membran Plasma Dan Transfor AktifDokumen9 halamanMakalah Tentang Membran Plasma Dan Transfor AktifRahayuBelum ada peringkat
- WRD, Referat, GNAPSDokumen29 halamanWRD, Referat, GNAPSAce ZamzamBelum ada peringkat
- Hasil PBL EdemaDokumen26 halamanHasil PBL EdemaWinda Diah NugraheniBelum ada peringkat
- PDF Laporan Kasus Perdarahan Akibat Defisiensi Vitamin K CompressDokumen1 halamanPDF Laporan Kasus Perdarahan Akibat Defisiensi Vitamin K CompressFahmi SuhandinataBelum ada peringkat
- Laporan Kasus yDokumen35 halamanLaporan Kasus yFahmi SuhandinataBelum ada peringkat
- Mekanisme Pengumpulan Artikel Pada Open Journal SystemDokumen2 halamanMekanisme Pengumpulan Artikel Pada Open Journal SystemFahmi SuhandinataBelum ada peringkat
- Ipe 1Dokumen4 halamanIpe 1Fahmi SuhandinataBelum ada peringkat
- Full SHDokumen14 halamanFull SHFahmi SuhandinataBelum ada peringkat
- LAPORAN KASUsDokumen33 halamanLAPORAN KASUsFahmi SuhandinataBelum ada peringkat
- Keompok A Higiene FixDokumen31 halamanKeompok A Higiene FixFahmi SuhandinataBelum ada peringkat
- Tatalaksana SHDokumen5 halamanTatalaksana SHFahmi SuhandinataBelum ada peringkat
- DM Dalam KehamilanDokumen32 halamanDM Dalam KehamilanDhiastikaBelum ada peringkat
- SCI - FahmiDokumen15 halamanSCI - FahmiFahmi SuhandinataBelum ada peringkat
- TX SHDokumen4 halamanTX SHFahmi SuhandinataBelum ada peringkat
- Lapsus SNDokumen35 halamanLapsus SNFahmi SuhandinataBelum ada peringkat
- Asam NukleatDokumen61 halamanAsam NukleatLianSiahaanBelum ada peringkat
- Ekspresi Gen Bu AndinDokumen51 halamanEkspresi Gen Bu Andinanida septiBelum ada peringkat
- Albumin Ikan Gabus Sebagai Anti Inflamasi Pada Sinusitis KronisDokumen18 halamanAlbumin Ikan Gabus Sebagai Anti Inflamasi Pada Sinusitis KronisFahmi SuhandinataBelum ada peringkat
- DPMDokumen1 halamanDPMFahmi SuhandinataBelum ada peringkat
- Saluran Nafas Atas 1 Rhinitis Alergi Dan AsmaDokumen12 halamanSaluran Nafas Atas 1 Rhinitis Alergi Dan AsmaFahmi SuhandinataBelum ada peringkat
- Mekanisme Remodeling Jalan Napas Atas Dan BawahDokumen17 halamanMekanisme Remodeling Jalan Napas Atas Dan BawahFahmi SuhandinataBelum ada peringkat
- Tinjauan Artikel Albumin Sebagai Obat Efek Biologis Albumin Tidak Terkait Dengan Tekanan OnkotikDokumen8 halamanTinjauan Artikel Albumin Sebagai Obat Efek Biologis Albumin Tidak Terkait Dengan Tekanan OnkotikFahmi SuhandinataBelum ada peringkat
- Kelompok 11 B Minggu 3Dokumen28 halamanKelompok 11 B Minggu 3Fahmi SuhandinataBelum ada peringkat
- Kanggo TM, GO Lan MASTADokumen4 halamanKanggo TM, GO Lan MASTAFahmi SuhandinataBelum ada peringkat
- Persentasi JurnalDokumen36 halamanPersentasi JurnalFahmi SuhandinataBelum ada peringkat
- ReferatDokumen25 halamanReferatFahmi SuhandinataBelum ada peringkat
- Imm SosmaDokumen10 halamanImm SosmaFahmi SuhandinataBelum ada peringkat
- Referat Xeroftalmia DR JannesDokumen38 halamanReferat Xeroftalmia DR JannesRiduan Adoro Lumban Gaol100% (2)
- Pemeriksaan Trikomoniasis, PPTDokumen22 halamanPemeriksaan Trikomoniasis, PPTFahmi Suhandinata100% (1)
- Lupus Erythematosus DiscoidDokumen18 halamanLupus Erythematosus DiscoiddgumelarBelum ada peringkat
- Anatomi Dan Fisiologi Sinus para AsalDokumen6 halamanAnatomi Dan Fisiologi Sinus para AsalFahmi SuhandinataBelum ada peringkat
- Case Report Tonsilitis KronisDokumen27 halamanCase Report Tonsilitis KronisFahmi SuhandinataBelum ada peringkat