Anda mungkin juga menyukai
- Peran Cahaya Dalam Tanaman - FotomorfogenesisDokumen27 halamanPeran Cahaya Dalam Tanaman - FotomorfogenesisUmi TrisnaningsihBelum ada peringkat
- NCCCCCCCCCCCDokumen17 halamanNCCCCCCCCCCCaliza khumairaBelum ada peringkat
- RESPON FOTODokumen55 halamanRESPON FOTOZakhruf As-salaamahBelum ada peringkat
- Fotomorfogenesis: Mengerti Respon Tanaman terhadap CahayaDokumen20 halamanFotomorfogenesis: Mengerti Respon Tanaman terhadap CahayaRiri Rahmadani PutriBelum ada peringkat
- Respon TumbuhanDokumen15 halamanRespon TumbuhanMohammad ShohibBelum ada peringkat
- Bab 22 FullDokumen21 halamanBab 22 FullHaryatin Nurul AfifahBelum ada peringkat
- Fotomorfogenesis Kelompok 4Dokumen49 halamanFotomorfogenesis Kelompok 4zuhrotul mufidahBelum ada peringkat
- FOTOMORFOGENESISDokumen4 halamanFOTOMORFOGENESISAnisah MahmudahBelum ada peringkat
- Fitokrom Paling Berpengaruh Pada Proses PhotomofogenesisDokumen3 halamanFitokrom Paling Berpengaruh Pada Proses PhotomofogenesisIdha Nur'ainiBelum ada peringkat
- P2-Plant Physiolo-Photomorphogenesis-g01.en - IdDokumen18 halamanP2-Plant Physiolo-Photomorphogenesis-g01.en - IdMaharani Ayu Safitri 118Belum ada peringkat
- Gabungn Fistum PresentasiDokumen20 halamanGabungn Fistum PresentasiBagoes SatrioBelum ada peringkat
- FOTOMORFOGENESIS DAN PENERIMA CAHAYADokumen15 halamanFOTOMORFOGENESIS DAN PENERIMA CAHAYAPrasetyaBelum ada peringkat
- Fisiologi TumbuhanDokumen100 halamanFisiologi Tumbuhanike nursantiBelum ada peringkat
- Paper Fistum FotomorfogenesisDokumen15 halamanPaper Fistum FotomorfogenesisGalang Syahya75% (4)
- Risma Amalya A - 200341617288 - OFFR B - POSTTEST FOTOMORFOGENESIS DAN FITOKROMDokumen4 halamanRisma Amalya A - 200341617288 - OFFR B - POSTTEST FOTOMORFOGENESIS DAN FITOKROMRisma AmalyaBelum ada peringkat
- Makalah FITOKROMDokumen13 halamanMakalah FITOKROMibhe123100% (1)
- Pigmen FotosintetikDokumen15 halamanPigmen FotosintetikRizky AlfarizyBelum ada peringkat
- Fotomorfogenesis-WPS OfficeDokumen6 halamanFotomorfogenesis-WPS OfficeAditiya NurrahmaBelum ada peringkat
- Kel 5Dokumen21 halamanKel 5JefonBelum ada peringkat
- Materi 9-Phytochrome-Light Control PDFDokumen33 halamanMateri 9-Phytochrome-Light Control PDFkontoladoBelum ada peringkat
- Fitokrom dan Pengaturan TumbuhanDokumen9 halamanFitokrom dan Pengaturan TumbuhanHepzhy SienaBelum ada peringkat
- Makalah Fotomorfogenesis PDFDokumen17 halamanMakalah Fotomorfogenesis PDFFatkurBelum ada peringkat
- FOTO MORFOGENESISDokumen17 halamanFOTO MORFOGENESISSanti ErynaBelum ada peringkat
- Peran Fitokrom Pada TumbuhanDokumen2 halamanPeran Fitokrom Pada TumbuhanHaafizh Sejati100% (1)
- FOTOREGULASIDokumen10 halamanFOTOREGULASIArdianBelum ada peringkat
- FOTOTROPISMEDokumen20 halamanFOTOTROPISMEarwiniBelum ada peringkat
- Fotosintesis PDFDokumen18 halamanFotosintesis PDFSepha Ronaldo100% (1)
- Empat Kelompok Fotoreseptor Pada TumbuhanDokumen1 halamanEmpat Kelompok Fotoreseptor Pada TumbuhanRiska May HabibiBelum ada peringkat
- DIT - M24 FotomorfogenisisDokumen12 halamanDIT - M24 FotomorfogenisisUtari KusumadewiBelum ada peringkat
- Tumbuhan CahayaDokumen12 halamanTumbuhan Cahayaanggraini putri utamiBelum ada peringkat
- Terjemahan Chapter 13Dokumen77 halamanTerjemahan Chapter 13kiranaBelum ada peringkat
- Terjemahan Chapter 13 MikrobiologiDokumen77 halamanTerjemahan Chapter 13 MikrobiologiLastri TriBelum ada peringkat
- MAKALAH FISIOLOGI TANAMAN "Phytochrome and Light Control of Plant Development"Dokumen13 halamanMAKALAH FISIOLOGI TANAMAN "Phytochrome and Light Control of Plant Development"towah yudiBelum ada peringkat
- Fotomorfogenesis dan FitokromDokumen19 halamanFotomorfogenesis dan FitokromnadyasuwayviaBelum ada peringkat
- FotomorfogenesisDokumen11 halamanFotomorfogenesisIjal AusiBelum ada peringkat
- Klorofil SpektroskopiDokumen11 halamanKlorofil SpektroskopiBela SuryaBelum ada peringkat
- Tugas Fistum LuckyDokumen1 halamanTugas Fistum LuckyLucky EkaBelum ada peringkat
- Fotografi CahayaDokumen8 halamanFotografi CahayaRisqy Amalia FarhanBelum ada peringkat
- PENGARUH KUALITAS RADIASI MATAHARI TERHADAP PERTUMBUHAN TANAMANDokumen4 halamanPENGARUH KUALITAS RADIASI MATAHARI TERHADAP PERTUMBUHAN TANAMANSanta TheresiasitinjakBelum ada peringkat
- Resume Fotosintesis 1Dokumen11 halamanResume Fotosintesis 1FnalifahBelum ada peringkat
- KADAR KLOROFILDokumen16 halamanKADAR KLOROFILSophan HadieBelum ada peringkat
- Rescued DocumentDokumen15 halamanRescued DocumentTantan HadiansyahBelum ada peringkat
- Laporan PigmenDokumen5 halamanLaporan PigmenAmila SholihatBelum ada peringkat
- Laporan Praktikum KlorofilDokumen4 halamanLaporan Praktikum KlorofilAnggi KlaritaBelum ada peringkat
- FitokromDokumen4 halamanFitokromEga Aris SenaBelum ada peringkat
- FOTODokumen6 halamanFOTOMisti17Belum ada peringkat
- FOTOSINTESIS DAN ASIMILASI PADA TUMBUHAN - KLMPK 4 Topik 6Dokumen14 halamanFOTOSINTESIS DAN ASIMILASI PADA TUMBUHAN - KLMPK 4 Topik 6itaBelum ada peringkat
- Fotomorfogenesis Dan FotoperiodismeDokumen20 halamanFotomorfogenesis Dan FotoperiodismefatehahwatiBelum ada peringkat
- Script BioDokumen3 halamanScript BioSinta Official12Belum ada peringkat
- Tugas 3 FotosintesisDokumen8 halamanTugas 3 FotosintesisUlviyardes DondanBelum ada peringkat
- Review Jurnal FISTAN - 632101040004 - Sofi Soraya Mulandani RahmatDokumen3 halamanReview Jurnal FISTAN - 632101040004 - Sofi Soraya Mulandani RahmatSofi SorayaBelum ada peringkat
- FotosintesisDokumen22 halamanFotosintesisArlinda Puspita SariBelum ada peringkat
- Materi 04 - Ekologi Tanaman HortikulturaDokumen25 halamanMateri 04 - Ekologi Tanaman HortikulturaPrisca 1100% (2)
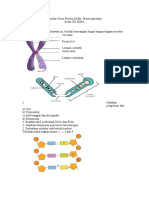
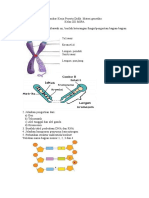
- Lembar Kerja Peserta Didik Materi Genetika Klp-1Dokumen1 halamanLembar Kerja Peserta Didik Materi Genetika Klp-1bqnovi14Belum ada peringkat
- Jurnal Harian Pekan 1-2Dokumen8 halamanJurnal Harian Pekan 1-2bqnovi14Belum ada peringkat
- GENETIKADokumen3 halamanGENETIKAbqnovi14Belum ada peringkat
- Jurnal Harian Pekan 2-1Dokumen7 halamanJurnal Harian Pekan 2-1bqnovi14Belum ada peringkat
- Lembar Kerja Peserta Didik Materi Genetika Klp-2Dokumen1 halamanLembar Kerja Peserta Didik Materi Genetika Klp-2bqnovi14Belum ada peringkat
- 2 Kompetensi - Inti - Dan - Komptensi - Dasar - Biologi - Sma - 37 - 2018Dokumen6 halaman2 Kompetensi - Inti - Dan - Komptensi - Dasar - Biologi - Sma - 37 - 2018Chika AnzilaBelum ada peringkat
- RPP Biologi VirusDokumen7 halamanRPP Biologi Virusbqnovi14Belum ada peringkat
- ALAT PENILAIANDokumen8 halamanALAT PENILAIANbqnovi14Belum ada peringkat
- Kemuhammadiyahan dan Pembaruan IslamDokumen3 halamanKemuhammadiyahan dan Pembaruan Islambqnovi14Belum ada peringkat
- KeanekaragamanHayatiDokumen48 halamanKeanekaragamanHayatibqnovi14Belum ada peringkat
- Bq. Novi Aprilia - Strategi Pembelajaran BiologiDokumen6 halamanBq. Novi Aprilia - Strategi Pembelajaran Biologibqnovi14Belum ada peringkat
- LangkerDokumen3 halamanLangkerbqnovi14Belum ada peringkat
- Bq. Novi Aprilia (E1A019021)Dokumen93 halamanBq. Novi Aprilia (E1A019021)bqnovi14Belum ada peringkat
- MANGROVEDokumen9 halamanMANGROVEbqnovi14Belum ada peringkat
- Cover Laporan PraktikumDokumen4 halamanCover Laporan Praktikumbqnovi14Belum ada peringkat
- MANGROVEDokumen9 halamanMANGROVEbqnovi14Belum ada peringkat