Anda mungkin juga menyukai
- Biosel Kelompok 6Dokumen11 halamanBiosel Kelompok 6NoviBelum ada peringkat
- Makalah Genetika Kel. 3Dokumen34 halamanMakalah Genetika Kel. 3mutiara indah lestariBelum ada peringkat
- Makalah Biomedik Sintesis ProteinDokumen21 halamanMakalah Biomedik Sintesis ProteinRatu Syamsiah Nila KencanaBelum ada peringkat
- Transkripsi Translasi Dan Pematangan MRN (Mba Yul) - 1Dokumen16 halamanTranskripsi Translasi Dan Pematangan MRN (Mba Yul) - 1Bella TrixBelum ada peringkat
- Sentral DogmaDokumen19 halamanSentral DogmaNadiyaBelum ada peringkat
- Makalah Materi GenetikDokumen25 halamanMakalah Materi GenetikDesty Febriani SosibongBelum ada peringkat
- TUGAS MAKALAH Sintesis ProteinDokumen12 halamanTUGAS MAKALAH Sintesis Proteinjanescaginting100% (1)
- Tugs Prof. EtiDokumen20 halamanTugs Prof. EtiUntoro Heri saputroBelum ada peringkat
- MAKALAH BIOLOGI Dhea AnandaDokumen12 halamanMAKALAH BIOLOGI Dhea AnandaDhea AnandaBelum ada peringkat
- UAS MODUL Transkripsi Dan Kontrol Dari Ekspresi Gen - Kelompok 6Dokumen20 halamanUAS MODUL Transkripsi Dan Kontrol Dari Ekspresi Gen - Kelompok 6Hesti ekaBelum ada peringkat
- Makalah Replikasi Transkripsi Dna - Biosel'Dokumen16 halamanMakalah Replikasi Transkripsi Dna - Biosel'tharaBelum ada peringkat
- Replikasi Dna Dan TranskripsiDokumen6 halamanReplikasi Dna Dan Transkripsiandini lestariBelum ada peringkat
- KelasA Tugas1A ZakkaFadhilah 150510200107Dokumen6 halamanKelasA Tugas1A ZakkaFadhilah 150510200107ZAKKA FADHILAHBelum ada peringkat
- PR - TP Hematologi Dr. ShofiaDokumen5 halamanPR - TP Hematologi Dr. ShofiashofiawmBelum ada peringkat
- Translasi BioselDokumen17 halamanTranslasi BioselYuni SetiyowatiBelum ada peringkat
- Kata PengantarDokumen10 halamanKata PengantarGugun gumilarBelum ada peringkat
- MAKALAH SINTESIS PROTEIN MunaDokumen9 halamanMAKALAH SINTESIS PROTEIN MunaCut SariBelum ada peringkat
- Sintesis ProteinDokumen19 halamanSintesis ProteindelaBelum ada peringkat
- G30122033 - Stabilitas Asam Nukleat Dan Sistem Ekspresi GenDokumen13 halamanG30122033 - Stabilitas Asam Nukleat Dan Sistem Ekspresi GenNi komang ayu PuspitasariBelum ada peringkat
- Makalah Sintesis Protein HanifahDokumen9 halamanMakalah Sintesis Protein HanifahCut SariBelum ada peringkat
- Bioinformatika Regita Ardhi A. 162210101031Dokumen196 halamanBioinformatika Regita Ardhi A. 162210101031Yani Putri RBelum ada peringkat
- Makalah Sintesis ProteinDokumen10 halamanMakalah Sintesis ProteinRE Angkat100% (1)
- Biosel Replikasi DnaDokumen18 halamanBiosel Replikasi Dnayolanda bupuBelum ada peringkat
- Azizah Nur AR - N012181023 - Transkripsi Pada EukariotikDokumen27 halamanAzizah Nur AR - N012181023 - Transkripsi Pada EukariotikAnidraqif HastinaBelum ada peringkat
- Makala HDokumen11 halamanMakala H22.Rifaldi RahmawanBelum ada peringkat
- Makalah Mekanisme Transkripsi Pada ProkariotDokumen42 halamanMakalah Mekanisme Transkripsi Pada ProkariotchiciangrniBelum ada peringkat
- Genetika TiyoDokumen11 halamanGenetika Tiyo9yft6sb48cBelum ada peringkat
- Sintesis Protein Pada Prokariot Dan EukariotDokumen31 halamanSintesis Protein Pada Prokariot Dan EukariotNi Komang Puja Pertiwi / 30Belum ada peringkat
- Makalah Replikasi Transkripsi Dna - Biosel'Dokumen16 halamanMakalah Replikasi Transkripsi Dna - Biosel'Yoki Robiyanto AlfathBelum ada peringkat
- Makalah Biologi Sel Ekspresi Gen Dan Mekanisme PengendalianDokumen16 halamanMakalah Biologi Sel Ekspresi Gen Dan Mekanisme PengendalianPutri MeilaniBelum ada peringkat
- Biosintesis Dna Dan Rna Hewan-1Dokumen22 halamanBiosintesis Dna Dan Rna Hewan-1Sri qumahira IraBelum ada peringkat
- Makalah Dogma Sentral Kel. 2Dokumen21 halamanMakalah Dogma Sentral Kel. 2qiungbBelum ada peringkat
- Makalah Sintesis ProteinDokumen18 halamanMakalah Sintesis ProteinNurhayati R Baruadi100% (1)
- Sintesa ProteinDokumen15 halamanSintesa ProteinMega ApryantiBelum ada peringkat
- Sintesis ProteinDokumen21 halamanSintesis ProteinFa ArdianaBelum ada peringkat
- Aliran Informasi GenetikDokumen11 halamanAliran Informasi GenetikPoppy ZuhBelum ada peringkat
- Kelas A-Tugas 1b-Kelompok 2Dokumen13 halamanKelas A-Tugas 1b-Kelompok 2NR12Belum ada peringkat
- Makalah GenetikDokumen29 halamanMakalah GenetikFina Triana SambiteBelum ada peringkat
- Translasi Pro, EukarDokumen14 halamanTranslasi Pro, EukarFensi AndraBelum ada peringkat
- Rizki Samsul Kurnia 1Dokumen13 halamanRizki Samsul Kurnia 1Bem LarukuBelum ada peringkat
- Makalah Sintesis ProteinDokumen11 halamanMakalah Sintesis ProteinEka Putri LiyaniBelum ada peringkat
- Tugas Biomol - GenDokumen9 halamanTugas Biomol - GenArendraBelum ada peringkat
- Makalah Biologi Molekuler Kelompok 2Dokumen17 halamanMakalah Biologi Molekuler Kelompok 2Fauziah SafitriBelum ada peringkat
- Makalah Sintesa ProteinDokumen22 halamanMakalah Sintesa ProteinerinnurbadriahBelum ada peringkat
- Makalah Sintesis Protein (NUR ANNISA)Dokumen18 halamanMakalah Sintesis Protein (NUR ANNISA)Nur AnnisaBelum ada peringkat
- Processing RnaDokumen13 halamanProcessing RnaMeilana Sapta DBelum ada peringkat
- Biomol Dna, Gen, GenomDokumen16 halamanBiomol Dna, Gen, GenomMarolop NapitupuluBelum ada peringkat
- TUGAS Biomol NurwiatiDokumen4 halamanTUGAS Biomol NurwiatiNurwiatiBelum ada peringkat
- Sintesis Protein (Ekspresi Gen)Dokumen8 halamanSintesis Protein (Ekspresi Gen)sylfiana harvaniBelum ada peringkat
- Aliran Informasi GenetikDokumen11 halamanAliran Informasi GenetikSepti AndrianiBelum ada peringkat
- MAKALAH TEKBIOMOL KEL 1 PerbaruhiDokumen17 halamanMAKALAH TEKBIOMOL KEL 1 PerbaruhiIlu Sulfihat ParawansaBelum ada peringkat
- UTS BiotekDokumen3 halamanUTS BiotekAbyan FarhandhityaBelum ada peringkat
- Sintesis ProteinDokumen11 halamanSintesis ProteinKiki Nur AzizahBelum ada peringkat
- 37 - Raudha Isminiarti Izza - Uas - BiokimDokumen3 halaman37 - Raudha Isminiarti Izza - Uas - BiokimYhessie OctaviaBelum ada peringkat
- SISTEM PROTEIN (IBD) YanaDokumen10 halamanSISTEM PROTEIN (IBD) YanaYana TalapessyBelum ada peringkat
- Laporan PraktikumDokumen13 halamanLaporan PraktikumSop IaBelum ada peringkat
- Kelompok 4 - Suksesi EkologiDokumen30 halamanKelompok 4 - Suksesi EkologiShabrina LieBelum ada peringkat
- Manajemen BisnisDokumen31 halamanManajemen BisnisShabrina LieBelum ada peringkat
- Prosedur Kerja Dan Pembahasan PCQDokumen5 halamanProsedur Kerja Dan Pembahasan PCQShabrina LieBelum ada peringkat
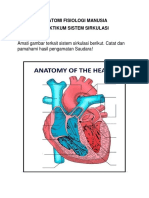
- Praktikum Sistem SirkulasiDokumen4 halamanPraktikum Sistem SirkulasiShabrina LieBelum ada peringkat
- Hasil Diskusi KH Kelompok 1 Off B 2019Dokumen6 halamanHasil Diskusi KH Kelompok 1 Off B 2019Shabrina LieBelum ada peringkat
- Modul KWUDokumen98 halamanModul KWURiva NoviaBelum ada peringkat
- Tata Cara OnlineDokumen28 halamanTata Cara OnlineShabrina LieBelum ada peringkat
- Kelompok 4 - Suksesi EkologiDokumen30 halamanKelompok 4 - Suksesi EkologiShabrina LieBelum ada peringkat
- Kelompok 4 - Mikrobiologi (Imunologi Dan Flora Normal Tubuh Manusia)Dokumen38 halamanKelompok 4 - Mikrobiologi (Imunologi Dan Flora Normal Tubuh Manusia)Shabrina LieBelum ada peringkat
- Ekologi ShabyDokumen4 halamanEkologi ShabyShabrina LieBelum ada peringkat
- Kelompok 3 EkologiDokumen22 halamanKelompok 3 EkologiShabrina LieBelum ada peringkat
- Kelompok 10 Makalah MetamorphosisDokumen19 halamanKelompok 10 Makalah MetamorphosisShabrina LieBelum ada peringkat
- Ecology PPT Suksesi EkologiDokumen56 halamanEcology PPT Suksesi EkologiShabrina LieBelum ada peringkat
- Tumbuhan LumutDokumen29 halamanTumbuhan LumutShabrina Lie100% (1)
- Format ANTISDokumen9 halamanFormat ANTISShabrina LieBelum ada peringkat
- Laporan Praktikum Mikrobiologi Shabrina Laili 190341621609Dokumen23 halamanLaporan Praktikum Mikrobiologi Shabrina Laili 190341621609Shabrina LieBelum ada peringkat
- Tumbuhan LumutDokumen29 halamanTumbuhan LumutShabrina Lie100% (1)
- PKN PartisipasiPolitikDemorasi - SpiDokumen15 halamanPKN PartisipasiPolitikDemorasi - SpiShabrina LieBelum ada peringkat
- Yuk Di BacaDokumen32 halamanYuk Di BacaAzisBelum ada peringkat
- Anatomi Sistem RangkaDokumen28 halamanAnatomi Sistem RangkaaristaBelum ada peringkat
- SE 10 Tahun 2021 Petunjuk Pelaksana Perjalanan Orang Dalam Negeri Dengan Transportasi Udara Dalam Masa Pandemi Covid-19Dokumen5 halamanSE 10 Tahun 2021 Petunjuk Pelaksana Perjalanan Orang Dalam Negeri Dengan Transportasi Udara Dalam Masa Pandemi Covid-19Indra Komara NugrahaBelum ada peringkat
- krs190341621609 20202Dokumen1 halamankrs190341621609 20202Shabrina LieBelum ada peringkat
- B Sunderland 1001Dokumen194 halamanB Sunderland 1001Shabrina LieBelum ada peringkat
- Praktikum 5 PCQ Edited 2021Dokumen8 halamanPraktikum 5 PCQ Edited 2021Shabrina LieBelum ada peringkat
- Petunjuk Praktikum-Rangka Dan OtotDokumen32 halamanPetunjuk Praktikum-Rangka Dan OtotShabrina LieBelum ada peringkat
- Identifikasi Filum Mollusca (Gastropoda) Di Perairan Palipi Soreang Kec. BaDokumen103 halamanIdentifikasi Filum Mollusca (Gastropoda) Di Perairan Palipi Soreang Kec. BaHikmah AsdinBelum ada peringkat
- Turunan Mesoderm Urogenital Limb 2014Dokumen18 halamanTurunan Mesoderm Urogenital Limb 2014Fira SfrwpBelum ada peringkat
- Dinamika Pengelolaan Krisis Sumberdaya HDokumen24 halamanDinamika Pengelolaan Krisis Sumberdaya HShabrina LieBelum ada peringkat
- 2020 RPS Dan SAP Belajar Dan Pembelajaran New Tim MKUDokumen5 halaman2020 RPS Dan SAP Belajar Dan Pembelajaran New Tim MKUSalvia Novanni ChaerunissaBelum ada peringkat
- Ponidin-Djj-Mpqlpf, Hmyoyw-Srg-Flight - OriginatingDokumen5 halamanPonidin-Djj-Mpqlpf, Hmyoyw-Srg-Flight - OriginatingPonidinFikomBelum ada peringkat