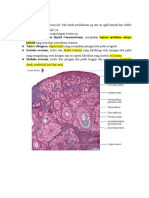
Anda mungkin juga menyukai
- Resume 2 - Regulasi Ekspresi Gen Pada EukariotikDokumen10 halamanResume 2 - Regulasi Ekspresi Gen Pada EukariotikNindis Pristya100% (1)
- Biologi - TO Akbar 1 Sanggar EdukasiDokumen6 halamanBiologi - TO Akbar 1 Sanggar EdukasiAnita BudiBelum ada peringkat
- Evolusi Tumbuhan BerbijiDokumen7 halamanEvolusi Tumbuhan BerbijiAri Yuni PriyaniBelum ada peringkat
- Uji MolishDokumen4 halamanUji MolishifaBelum ada peringkat
- Struktur Anatomi Buah Dan Biji Sirsak (AnnonaDokumen2 halamanStruktur Anatomi Buah Dan Biji Sirsak (AnnonaKhevakwaBelum ada peringkat
- Pertumbuhan Dan AnDokumen46 halamanPertumbuhan Dan AnYusi Listiawati100% (1)
- PENDAHULUA1Dokumen15 halamanPENDAHULUA1ifan_elfananyBelum ada peringkat
- Analisis Sifat KarbohidratDokumen24 halamanAnalisis Sifat KarbohidratResa Indriawan100% (1)
- Soal Siklus SelDokumen2 halamanSoal Siklus SelRaja Fauzan FahleviBelum ada peringkat
- Ciri-Ciri, Reproduksi, Dan Klasifikasi Tumbuhan Lumut (Bryophyta)Dokumen6 halamanCiri-Ciri, Reproduksi, Dan Klasifikasi Tumbuhan Lumut (Bryophyta)Lilik Niko Dwijayanto100% (2)
- Glikolisis Dari SukrosaDokumen6 halamanGlikolisis Dari SukrosaNugroho Azis Wahyu PrasetyoBelum ada peringkat
- Peran Biosistematika Dan ContohnyaDokumen3 halamanPeran Biosistematika Dan ContohnyaFaiz MuhammadBelum ada peringkat
- Pembahasan MnO2 Dan H2O2Dokumen2 halamanPembahasan MnO2 Dan H2O2Dinda KhumayirohBelum ada peringkat
- Struktur Jaringan OvariumDokumen6 halamanStruktur Jaringan OvariumFarah Fadhilah Rosyadi100% (1)
- Pengertian Metagenesis Dan Penjelasan Proses Metagenesis (Lengkap)Dokumen9 halamanPengertian Metagenesis Dan Penjelasan Proses Metagenesis (Lengkap)Bartolomeus LelawayangBelum ada peringkat
- Reproduksi Sel, Hewan, Dan TumbuhanDokumen6 halamanReproduksi Sel, Hewan, Dan TumbuhanFaishal Aji ZulmiBelum ada peringkat
- Praktikum 2 BocryptDokumen9 halamanPraktikum 2 BocryptHari Muhammad FirdausBelum ada peringkat
- Ringkasan Embriologi Perbandingan - Kelompok 3 - PBA 2018Dokumen1 halamanRingkasan Embriologi Perbandingan - Kelompok 3 - PBA 2018murid rumahanBelum ada peringkat
- Rangkuman Struktur Dan Fungsi Jaringan TumbuhanDokumen3 halamanRangkuman Struktur Dan Fungsi Jaringan Tumbuhanirma indanaBelum ada peringkat
- Tugas SPH 1 Sistem Sirkulasi Karman NusiDokumen3 halamanTugas SPH 1 Sistem Sirkulasi Karman NusiKarman NusiBelum ada peringkat
- Klasifikasi of FungiDokumen13 halamanKlasifikasi of FungiFatimahAzZahraBelum ada peringkat
- Alga CoklatDokumen13 halamanAlga CoklatLinda Sy HattaBelum ada peringkat
- Penuntun Biosistematik REVDokumen113 halamanPenuntun Biosistematik REVPutri AniBelum ada peringkat
- Jaringan EpitelDokumen8 halamanJaringan EpitelVandhaSalsabilaBelum ada peringkat
- Keanekaragaman Protista Dan FungiDokumen46 halamanKeanekaragaman Protista Dan FungiSeptiaBelum ada peringkat
- Daftar Pustaka LengkapDokumen3 halamanDaftar Pustaka LengkapaldaBelum ada peringkat
- Struktur JaringanDokumen19 halamanStruktur JaringanElva LuoBelum ada peringkat
- MutasiDokumen112 halamanMutasiZanty Rakhmania PutriBelum ada peringkat
- Hidrolisis ProteinDokumen36 halamanHidrolisis ProteinFebi rianiBelum ada peringkat
- Regulasi Lac OperonDokumen4 halamanRegulasi Lac OperonokutairieBelum ada peringkat
- Tracheophyta Fixed FixxxxxDokumen28 halamanTracheophyta Fixed FixxxxxLilis nurasiahBelum ada peringkat
- II. LUMUT, Paku, Dan AlgaDokumen47 halamanII. LUMUT, Paku, Dan AlgaMarissa HarahapBelum ada peringkat
- Laila Magfirah ReptilDokumen45 halamanLaila Magfirah Reptillaila magfirah mahmudBelum ada peringkat
- ZymogenDokumen10 halamanZymogenAnung Irawan PBelum ada peringkat
- Klasifikasi TumbuhanDokumen17 halamanKlasifikasi TumbuhanRaihan NafisBelum ada peringkat
- Materi Reaksi KorteksDokumen3 halamanMateri Reaksi KorteksyasaboyBelum ada peringkat
- Makalah Kelompok 2 - Ruangg Lingkup Evolusi - OFF C 2018Dokumen17 halamanMakalah Kelompok 2 - Ruangg Lingkup Evolusi - OFF C 2018Siti WidyawatiBelum ada peringkat
- GlioksisomDokumen7 halamanGlioksisomAster LienBelum ada peringkat
- Pengaruh PH Terhadap Kerja Enzim PtialinDokumen7 halamanPengaruh PH Terhadap Kerja Enzim PtialinAtika Anggraini100% (1)
- Laporan PraktikumDokumen10 halamanLaporan PraktikumMaylaBelum ada peringkat
- Pengamatan Zat Penyusun Tulang Ceker AyamDokumen7 halamanPengamatan Zat Penyusun Tulang Ceker AyamImaBelum ada peringkat
- OVARIUMDokumen7 halamanOVARIUMArizal AbdullahBelum ada peringkat
- 5 Pertanyaan Organogenesis Dan Neurolasi - Andro Chaesi Todoan Manullang - 2013091016Dokumen2 halaman5 Pertanyaan Organogenesis Dan Neurolasi - Andro Chaesi Todoan Manullang - 2013091016Andro Chaesi Todoan ManullangBelum ada peringkat
- Laporan Referensi KHDokumen2 halamanLaporan Referensi KHana saniaBelum ada peringkat
- 5 Kepunahan Massal Terbesar Di BUMIDokumen6 halaman5 Kepunahan Massal Terbesar Di BUMIEka S. SaputraBelum ada peringkat
- Biologi AkuDokumen7 halamanBiologi AkuXiePayah No Skill100% (1)
- Interaksi Biotik Dengan AbiotikDokumen5 halamanInteraksi Biotik Dengan AbiotikIsni Azizah MaulidaBelum ada peringkat
- Pengujian AmilumDokumen2 halamanPengujian AmilumDevy N. FadillaBelum ada peringkat
- Laporan Botani 7Dokumen12 halamanLaporan Botani 7ali lasoBelum ada peringkat
- Biofisika Membran SelDokumen121 halamanBiofisika Membran Selbrainace100% (1)
- SPH GonadDokumen9 halamanSPH GonadPrasetyaBelum ada peringkat
- Laporan Praktikum EnzimDokumen10 halamanLaporan Praktikum EnzimNabila MarthaBelum ada peringkat
- Persamaan Dan Perbedaan Sel Prokariotik Dan EukariotikDokumen1 halamanPersamaan Dan Perbedaan Sel Prokariotik Dan EukariotikFiier Rainn100% (1)
- Penerimaan Sinyal Dan Inisiasi TransduksiDokumen8 halamanPenerimaan Sinyal Dan Inisiasi TransduksiVica FirmaBelum ada peringkat
- Komunikasi SelDokumen30 halamanKomunikasi SelKelas Biologi2Belum ada peringkat
- Komunikasi SelDokumen29 halamanKomunikasi SelBiodi AnggrnBelum ada peringkat
- Tugas Kimed Ii - 1Dokumen11 halamanTugas Kimed Ii - 1ZainalSyahroniBelum ada peringkat
- Makalah Komunikasi Sel (Perfect Print)Dokumen14 halamanMakalah Komunikasi Sel (Perfect Print)Ermawati Erma100% (2)
- Second MassengerDokumen7 halamanSecond MassengerMa MedBelum ada peringkat
- Perdarahan Pasca EkstraksiDokumen3 halamanPerdarahan Pasca EkstraksiFildza HasnamudhiaBelum ada peringkat
- Bagian Ody - SPACE REGAINERDokumen7 halamanBagian Ody - SPACE REGAINERFildza HasnamudhiaBelum ada peringkat
- Ody - Klasifikasi MaloklusiDokumen10 halamanOdy - Klasifikasi MaloklusiFildza HasnamudhiaBelum ada peringkat
- Klasifikasi Maloklusi Angle& DeweyDokumen5 halamanKlasifikasi Maloklusi Angle& DeweyFildza Hasnamudhia100% (2)
- Try in Coping & Mahkota Dan Sementasi Tetap Mahkota TiruanDokumen12 halamanTry in Coping & Mahkota Dan Sementasi Tetap Mahkota TiruanFildza Hasnamudhia100% (3)
- Macam Macam Cengkeram Kawat Dan Logam Tuang Pada GTSL Akrilik Dan Kerangka LogamDokumen9 halamanMacam Macam Cengkeram Kawat Dan Logam Tuang Pada GTSL Akrilik Dan Kerangka LogamFildza HasnamudhiaBelum ada peringkat
- Penanganan Kegawatdaruratan Trauma OromaksilofasialDokumen22 halamanPenanganan Kegawatdaruratan Trauma OromaksilofasialFildza HasnamudhiaBelum ada peringkat
- Macam Macam Cengkeram Kawat Dan Logam Tuang Pada GTSL Akrilik Dan Kerangka LogamDokumen13 halamanMacam Macam Cengkeram Kawat Dan Logam Tuang Pada GTSL Akrilik Dan Kerangka LogamFildza Hasnamudhia100% (2)
- Anomali Gigi SK4!Dokumen10 halamanAnomali Gigi SK4!Fildza HasnamudhiaBelum ada peringkat
- Bahan Ody - SIKAP MENTAL PASIEN & Hubungan Kondisi Kesehatan DGN PerawatanDokumen10 halamanBahan Ody - SIKAP MENTAL PASIEN & Hubungan Kondisi Kesehatan DGN PerawatanFildza HasnamudhiaBelum ada peringkat
- Bahan Ody - Indikasi Kontraindikasi Gigi Tiruan Penuh Dan GTP TunggalDokumen3 halamanBahan Ody - Indikasi Kontraindikasi Gigi Tiruan Penuh Dan GTP TunggalFildza HasnamudhiaBelum ada peringkat
- Calon Calon Games JambaksosDokumen6 halamanCalon Calon Games JambaksosFildza HasnamudhiaBelum ada peringkat
- Kualitatif GajelasDokumen6 halamanKualitatif GajelasFildza HasnamudhiaBelum ada peringkat
- Bahan Ody - Indikasi Kontraindikasi MTP Dan MTP TunggalDokumen2 halamanBahan Ody - Indikasi Kontraindikasi MTP Dan MTP TunggalFildza HasnamudhiaBelum ada peringkat
- Bagian Ody - PatogenesisDokumen3 halamanBagian Ody - PatogenesisFildza HasnamudhiaBelum ada peringkat
- Bagian Ody - Pengaruh Menstruasi Ke Penyakit PeriodontalDokumen4 halamanBagian Ody - Pengaruh Menstruasi Ke Penyakit PeriodontalFildza HasnamudhiaBelum ada peringkat
- Bahan Ody - Klasifikasi Dan Dampak Kehilangan GigiDokumen5 halamanBahan Ody - Klasifikasi Dan Dampak Kehilangan GigiFildza HasnamudhiaBelum ada peringkat
- Merokok PD Penyakit PerioDokumen3 halamanMerokok PD Penyakit PerioFildza HasnamudhiaBelum ada peringkat
- Bagian Ody - Etiologi Dan PatogenesisDokumen6 halamanBagian Ody - Etiologi Dan PatogenesisFildza HasnamudhiaBelum ada peringkat
- Bahan Ody - Sistem Stomatognatik Dan Konsep OklusiDokumen8 halamanBahan Ody - Sistem Stomatognatik Dan Konsep OklusiFildza HasnamudhiaBelum ada peringkat
- Bagian Ody - Etiologi Abses PeriodontalDokumen2 halamanBagian Ody - Etiologi Abses PeriodontalFildza HasnamudhiaBelum ada peringkat
- Bagian Ody - Prinsip Mekanik Dalam Kontrol Kekuatan OrthodonticDokumen12 halamanBagian Ody - Prinsip Mekanik Dalam Kontrol Kekuatan OrthodonticFildza HasnamudhiaBelum ada peringkat
- STEINER - Lanjutan GhrenaDokumen4 halamanSTEINER - Lanjutan GhrenaFildza HasnamudhiaBelum ada peringkat
- Impaksi MakananDokumen6 halamanImpaksi MakananFildza HasnamudhiaBelum ada peringkat
- Bagian Ody - Liner BasisDokumen10 halamanBagian Ody - Liner BasisFildza HasnamudhiaBelum ada peringkat
- Bagian Ody - Etiologi Dan PatogenesisDokumen6 halamanBagian Ody - Etiologi Dan PatogenesisFildza HasnamudhiaBelum ada peringkat
- Bagian Ody - RadiografikDokumen8 halamanBagian Ody - RadiografikFildza HasnamudhiaBelum ada peringkat
- Proses Healing Dan Repair Setelah Terapi PeriodontalDokumen5 halamanProses Healing Dan Repair Setelah Terapi PeriodontalFildza HasnamudhiaBelum ada peringkat
- Bagian Ody - Liner BasisDokumen10 halamanBagian Ody - Liner BasisFildza HasnamudhiaBelum ada peringkat
- Ukuran PemusatanDokumen12 halamanUkuran PemusatanFildza HasnamudhiaBelum ada peringkat