Anda mungkin juga menyukai
- Pembahasan Hidrolisis MentegaDokumen1 halamanPembahasan Hidrolisis MentegaNoviaBelum ada peringkat
- Klasifikasi Lipid Menurut BloorDokumen3 halamanKlasifikasi Lipid Menurut BloorRizal Nur Huda100% (1)
- DetektorDokumen4 halamanDetektorYusuf Maulana MaulanaBelum ada peringkat
- Laporan GC PDFDokumen12 halamanLaporan GC PDFWahyu UswaBelum ada peringkat
- Lipid Dan Produk Alam Yang BerhubunganDokumen8 halamanLipid Dan Produk Alam Yang BerhubunganRiaBelum ada peringkat
- Albumin ProteinDokumen3 halamanAlbumin ProteinDita Dita PutriBelum ada peringkat
- Dye Reduction Test Abdulloh Khudry and Aprillia Fatma ElyDokumen5 halamanDye Reduction Test Abdulloh Khudry and Aprillia Fatma ElyApriLia Fatma Ely-ohorella100% (1)
- Dampak PengerasDokumen2 halamanDampak PengerasJafar SyidikBelum ada peringkat
- LipidDokumen6 halamanLipidDilla WahabBelum ada peringkat
- Glikogenesis Kelompok 2Dokumen10 halamanGlikogenesis Kelompok 2Laras BudyaningrumBelum ada peringkat
- FlavonoidDokumen22 halamanFlavonoiddiah triyunitaBelum ada peringkat
- Penentuan Kadar GlukosaDokumen12 halamanPenentuan Kadar GlukosaIngratsusi MarvianiBelum ada peringkat
- Isolasi Enzim Amilase Dan Uji Gula Reduksi Dari TaugeDokumen12 halamanIsolasi Enzim Amilase Dan Uji Gula Reduksi Dari TaugeLamtiarma PanjaitanBelum ada peringkat
- Kelompok 4 GLIKOGENESIS Hasil DiskusiDokumen3 halamanKelompok 4 GLIKOGENESIS Hasil Diskusisafira muliaBelum ada peringkat
- Reaksi Sintesis Grignard 1Dokumen5 halamanReaksi Sintesis Grignard 1TyawinataBelum ada peringkat
- Gambar Mikroskopis Kristal OsazonDokumen1 halamanGambar Mikroskopis Kristal OsazonAghnia Ahdanisa HaraBelum ada peringkat
- Soal Diskusi Boteknologi FarmasiDokumen2 halamanSoal Diskusi Boteknologi FarmasiHerlin AgustinaBelum ada peringkat
- Reaksi Uji Koagulasi PanasDokumen2 halamanReaksi Uji Koagulasi PanasAnonymous adhp66Belum ada peringkat
- Kel 5-Laporan Praktikum Farkol SkriningDokumen20 halamanKel 5-Laporan Praktikum Farkol SkriningRindi Irsan ArtomoBelum ada peringkat
- Analisis Karbohidrat Menggunakan Metode KolorimetriDokumen27 halamanAnalisis Karbohidrat Menggunakan Metode KolorimetriAnggita Rosiana PutriBelum ada peringkat
- Isolasi Dan Identifikasi Aspergillus SPP 3609c572Dokumen6 halamanIsolasi Dan Identifikasi Aspergillus SPP 3609c572Nesa Shifa OksandaBelum ada peringkat
- Bab I ArgentometriDokumen14 halamanBab I ArgentometriYuniPujiRahmawatiBelum ada peringkat
- Percobaan 5 (Isolasi Dan Pemurnian Awal)Dokumen32 halamanPercobaan 5 (Isolasi Dan Pemurnian Awal)estiBelum ada peringkat
- Dasar Teori Protein Dan LipidDokumen5 halamanDasar Teori Protein Dan LipidCamilla Tirani100% (1)
- Makalah Analisis Farmasi KCKTDokumen18 halamanMakalah Analisis Farmasi KCKTMelisa Klinik RiyenaBelum ada peringkat
- Metabolisme Asam NukleatDokumen28 halamanMetabolisme Asam NukleatLuthfi AffandhyBelum ada peringkat
- 1291 TP SpektrofotometerDokumen2 halaman1291 TP SpektrofotometerFadhil GhiyatsBelum ada peringkat
- 1 PBDokumen7 halaman1 PBrafael datuanBelum ada peringkat
- Review Jurnal Koo - Intan Cahya Aprillia - A28226771Dokumen4 halamanReview Jurnal Koo - Intan Cahya Aprillia - A28226771Intan Cahya AprilliaBelum ada peringkat
- Karakteristik HEWAN COBADokumen7 halamanKarakteristik HEWAN COBArehan_uchihaBelum ada peringkat
- Jurnal ModulDokumen9 halamanJurnal ModulMONIKA JELITA SIREGAR MONIKA JELITA SIREGARBelum ada peringkat
- Laporan Uji Koliform Pada Sediaan CairDokumen14 halamanLaporan Uji Koliform Pada Sediaan CairYossie Kharisma DewiBelum ada peringkat
- Reaksi KimiaDokumen22 halamanReaksi KimiaestinurulBelum ada peringkat
- Laporan Akhir Praktikum BiokimiaDokumen22 halamanLaporan Akhir Praktikum BiokimiaSholihah AmrinaBelum ada peringkat
- Laporan Praktikum Analisis Farmasi KGDokumen9 halamanLaporan Praktikum Analisis Farmasi KGsherlyBelum ada peringkat
- Sintesis KolesterolDokumen1 halamanSintesis KolesterolDeliBelum ada peringkat
- Biosintesis Asam Lemak JenuhDokumen4 halamanBiosintesis Asam Lemak Jenuhanon_217948194Belum ada peringkat
- High Performance Liquid ChromatographyDokumen24 halamanHigh Performance Liquid ChromatographyMuhammad Aldila SatriaBelum ada peringkat
- Analisis ProteinDokumen17 halamanAnalisis ProteinAyuBelum ada peringkat
- Pemanfaatan Ekstrak Enzim Papain Dan Pemanfaatannya Untuk Isolasi DnaDokumen14 halamanPemanfaatan Ekstrak Enzim Papain Dan Pemanfaatannya Untuk Isolasi DnaFauziyyah Diyah Anggita SariBelum ada peringkat
- Tinjauan PustakaDokumen10 halamanTinjauan PustakaKris Natalia ManihurukBelum ada peringkat
- Fungsi LipidDokumen2 halamanFungsi LipidAdlimatulPutriIlmiyahBelum ada peringkat
- Nujol MullDokumen9 halamanNujol MullRahmawati HamzainyBelum ada peringkat
- Pengaruh Logam Dan EdtaDokumen3 halamanPengaruh Logam Dan Edtawidya prastikaBelum ada peringkat
- MAKALAH - Farkol NkropsiDokumen11 halamanMAKALAH - Farkol NkropsiIsTi MasiversBelum ada peringkat
- Penentuan Kadar Iodium Dalam GaramDokumen21 halamanPenentuan Kadar Iodium Dalam GaramPutri Azizah100% (1)
- Total Plate CountDokumen21 halamanTotal Plate CountSahyudi Darma Asepti100% (1)
- Metode BradfordDokumen7 halamanMetode BradfordHerlani Tri WidhiastutiBelum ada peringkat
- Asam Karboksilat Dan TurunannyaDokumen49 halamanAsam Karboksilat Dan TurunannyaSerlyFebriantiBelum ada peringkat
- Tinjauan Pustaka KarbohidratDokumen4 halamanTinjauan Pustaka KarbohidratMeita Nazla AdilaBelum ada peringkat
- JURNAL KARBOHIDRAT. DeddyDokumen18 halamanJURNAL KARBOHIDRAT. DeddyMuhammadAtmanegaraBelum ada peringkat
- Percobaan 7 - 062119058 - Raden Thasya Puteri. RDokumen9 halamanPercobaan 7 - 062119058 - Raden Thasya Puteri. RThasya PuteriBelum ada peringkat
- Uji Kualitatif Kolesterol Kelompok A3Dokumen4 halamanUji Kualitatif Kolesterol Kelompok A3Dafa Sukma NugrahaBelum ada peringkat
- Laporan KBA Isolasi KarotenoidDokumen16 halamanLaporan KBA Isolasi KarotenoidarsyadBelum ada peringkat
- Pembahasan Uji Salkowski Kolestrol Dalam Kloroform FixDokumen3 halamanPembahasan Uji Salkowski Kolestrol Dalam Kloroform Fix21BELLAPERMATASARI kimiaBelum ada peringkat
- AgarosaDokumen3 halamanAgarosaMuaz Azhar100% (1)
- Biosintesis KolesterolDokumen6 halamanBiosintesis Kolesteroldellad16Belum ada peringkat
- BiosintesaDokumen9 halamanBiosintesaismi hayBelum ada peringkat
- Sintesis KolesterolDokumen3 halamanSintesis KolesterolImmanuel AgapaoBelum ada peringkat
- Biosintesis SteroidDokumen3 halamanBiosintesis SteroidgramatolinaBelum ada peringkat
- Nurul Muhlisa - LAPORAN LENGKAP PRAKTIKUM MIKOLOGIDokumen102 halamanNurul Muhlisa - LAPORAN LENGKAP PRAKTIKUM MIKOLOGILisa addanas TVBelum ada peringkat
- Laporan TNA - Tugas PBAKDokumen2 halamanLaporan TNA - Tugas PBAKLisa addanas TVBelum ada peringkat
- Tugas Matakuliah PBAKDokumen3 halamanTugas Matakuliah PBAKLisa addanas TVBelum ada peringkat
- Kelompok 10 Kewirausahaan - 1Dokumen26 halamanKelompok 10 Kewirausahaan - 1Lisa addanas TVBelum ada peringkat
- Tugas Pbak Nurul Muhlisa - Laporan TnaDokumen2 halamanTugas Pbak Nurul Muhlisa - Laporan TnaLisa addanas TVBelum ada peringkat
- Tugas MikologiDokumen9 halamanTugas MikologiLisa addanas TVBelum ada peringkat
- Tugas Imunohematologi PPT 9Dokumen2 halamanTugas Imunohematologi PPT 9Lisa addanas TVBelum ada peringkat
- Penjelasan KewirausahaanDokumen2 halamanPenjelasan KewirausahaanLisa addanas TVBelum ada peringkat
- Kelompok 3 Etika ProfesiDokumen11 halamanKelompok 3 Etika ProfesiLisa addanas TVBelum ada peringkat
- Escherichia Coli - Nurul MuhlisaDokumen21 halamanEscherichia Coli - Nurul MuhlisaAndi Ika PuspitasariBelum ada peringkat
- Laporan TNA - Tugas PBAKDokumen2 halamanLaporan TNA - Tugas PBAKLisa addanas TVBelum ada peringkat
- Kelompok Vi ProteusDokumen14 halamanKelompok Vi ProteusLisa addanas TVBelum ada peringkat
- 027 - Nurul Muhlisa - Tugas 123Dokumen18 halaman027 - Nurul Muhlisa - Tugas 123Lisa addanas TVBelum ada peringkat
- Streptococcus SP - Nurul MuhlisaDokumen20 halamanStreptococcus SP - Nurul MuhlisaLisa addanas TVBelum ada peringkat
- Klebsiella SP - Nurul MuhlisaDokumen20 halamanKlebsiella SP - Nurul MuhlisaLisa addanas TVBelum ada peringkat
- Staphylococcus SP - Nurul MuhlisaDokumen20 halamanStaphylococcus SP - Nurul MuhlisaLisa addanas TVBelum ada peringkat
- Plasmodium Knowlesi - Tugas Parasitologi Ii - Kelas ADokumen16 halamanPlasmodium Knowlesi - Tugas Parasitologi Ii - Kelas ALisa addanas TVBelum ada peringkat
- Proteus - Kelompok 6 - Bakteriologi Ii - Kelas ADokumen14 halamanProteus - Kelompok 6 - Bakteriologi Ii - Kelas ALisa addanas TVBelum ada peringkat
- Soal Kasus Biokimia Nurul MuhlisaDokumen20 halamanSoal Kasus Biokimia Nurul MuhlisaLisa addanas TVBelum ada peringkat
- 2.nurul Muhlisa - Tugas Imunoserologi I - Kelas ADokumen3 halaman2.nurul Muhlisa - Tugas Imunoserologi I - Kelas ALisa addanas TVBelum ada peringkat
- Media Pertumbuhan Bakteri - Nurul MuhlisaDokumen21 halamanMedia Pertumbuhan Bakteri - Nurul MuhlisaLisa addanas TVBelum ada peringkat
- 1.nurul Muhlisa - Tugas Imunoserologi I - Kelas ADokumen5 halaman1.nurul Muhlisa - Tugas Imunoserologi I - Kelas ALisa addanas TVBelum ada peringkat
- Mansonella OzzardiDokumen2 halamanMansonella OzzardiLisa addanas TVBelum ada peringkat
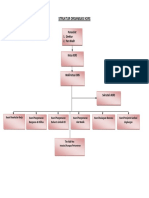
- Nurul Muhlisa - Struktur Organisasi K3 Laboratorium RSDokumen7 halamanNurul Muhlisa - Struktur Organisasi K3 Laboratorium RSLisa addanas TVBelum ada peringkat
- Intrumen Lab ParasitDokumen16 halamanIntrumen Lab ParasitLisa addanas TVBelum ada peringkat
- Makalah Instrumentasi Laboratorium ParasitologiDokumen26 halamanMakalah Instrumentasi Laboratorium ParasitologiLisa addanas TVBelum ada peringkat
- Makalah Brugya Malayi - Kelompok ViiDokumen15 halamanMakalah Brugya Malayi - Kelompok ViiLisa addanas TVBelum ada peringkat
- PRAKTIKUM IX (Nurul Muhlisa) - Isolasi Enzim BromelinDokumen4 halamanPRAKTIKUM IX (Nurul Muhlisa) - Isolasi Enzim BromelinLisa addanas TVBelum ada peringkat
- Biosafety Level (BSL) - K3Dokumen5 halamanBiosafety Level (BSL) - K3Lisa addanas TVBelum ada peringkat