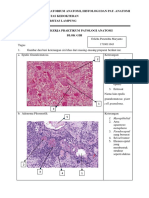
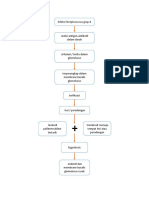
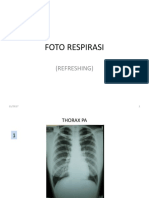
Anda mungkin juga menyukai
- Perkap Nomor 12 TTG Kedokteran KepolisianDokumen16 halamanPerkap Nomor 12 TTG Kedokteran KepolisianReinya DeviartiBelum ada peringkat
- Emfisema ParuDokumen23 halamanEmfisema ParuRises Christian0% (1)
- Komplikasi ITPDokumen17 halamanKomplikasi ITPFadillahulfa1099Belum ada peringkat
- Laporan HistologiDokumen13 halamanLaporan HistologifasyaBelum ada peringkat
- Wrap Up Skenario 2 FinalDokumen33 halamanWrap Up Skenario 2 Finalliki100% (1)
- SESAK NAPAS Blok Respirasi Skenario 3Dokumen44 halamanSESAK NAPAS Blok Respirasi Skenario 3Cory Dwi Rizki100% (1)
- Systemic Lupus Erythematosus (Sle)Dokumen28 halamanSystemic Lupus Erythematosus (Sle)Kristina Anita Meilani100% (1)
- Sistem Imun Adaptif 051020Dokumen59 halamanSistem Imun Adaptif 051020Andi SoewandiBelum ada peringkat
- Analisis Pengaruh Waktu Malam Hari Terhadap Munculnya Penyakit Asma Sebagai Dasar Preventif Asma NokturnalDokumen11 halamanAnalisis Pengaruh Waktu Malam Hari Terhadap Munculnya Penyakit Asma Sebagai Dasar Preventif Asma NokturnalIzzy VikratBelum ada peringkat
- 1.3 Morfologi ArbovirusDokumen10 halaman1.3 Morfologi Arbovirushafiz primadhyaBelum ada peringkat
- PorfiriaDokumen11 halamanPorfiriaFitrahBelum ada peringkat
- Sitokin & KemokinDokumen15 halamanSitokin & KemokinAhmat Rediansya PutraBelum ada peringkat
- Laporan PBL 2Dokumen14 halamanLaporan PBL 2wiwinwijayangsih100% (1)
- Patofisiologi HematuriaDokumen2 halamanPatofisiologi HematuriaRizky Fadhila ZunnuroinBelum ada peringkat
- Laporan Modul 2 Imunodefisiensi Kelompok 2Dokumen60 halamanLaporan Modul 2 Imunodefisiensi Kelompok 2ErmanW'theRachingMuhamadBelum ada peringkat
- Mikosis ParuDokumen24 halamanMikosis ParuEkwanto SBelum ada peringkat
- Mekanisme Pertahanan ParuDokumen3 halamanMekanisme Pertahanan ParuOpi Okvi KurniatiBelum ada peringkat
- Respon Imun SekunderDokumen1 halamanRespon Imun Sekunderdee.aira2955Belum ada peringkat
- Referat Trauma GinjalDokumen21 halamanReferat Trauma GinjalClinton SudjonoBelum ada peringkat
- Wrap Up Skenario 2 Blok Ginjal Dan Saluran Kemih "Kencing Sedikit"Dokumen22 halamanWrap Up Skenario 2 Blok Ginjal Dan Saluran Kemih "Kencing Sedikit"Ilham MahardikaBelum ada peringkat
- Lapkas - Simple PneumothoraxDokumen16 halamanLapkas - Simple PneumothoraxMohammad Hafidz Ramadhan100% (1)
- Batuk Berdahak Dan Sesak Nafas PneumoniaDokumen26 halamanBatuk Berdahak Dan Sesak Nafas PneumoniaRiska ULy100% (2)
- Laporan Nyeri DadaDokumen10 halamanLaporan Nyeri Dadayusrilkurniawan78Belum ada peringkat
- Pemeriksaan Penunjang PneumoniaDokumen2 halamanPemeriksaan Penunjang PneumoniaFatia ZulfaBelum ada peringkat
- Penyakit Jantung RematikDokumen13 halamanPenyakit Jantung RematikChristy AmandaBelum ada peringkat
- Laporan Skenario B Blok 16Dokumen51 halamanLaporan Skenario B Blok 16Catri Dwi Utari PramasariBelum ada peringkat
- Anatomi TonsilDokumen6 halamanAnatomi TonsildoooooiiiiiBelum ada peringkat
- PilekDokumen21 halamanPilekDeril RidwanBelum ada peringkat
- Neuritis VestibularisDokumen14 halamanNeuritis VestibularislipraaBelum ada peringkat
- Demam RematikDokumen17 halamanDemam RematikEva RosalinaBelum ada peringkat
- Tugas DD Nyeri SendiDokumen6 halamanTugas DD Nyeri SendiGinsha Audia KusumoBelum ada peringkat
- Drug Induced LupusDokumen8 halamanDrug Induced LupusSitaBelum ada peringkat
- LO Sinusitis EthmoidalisDokumen5 halamanLO Sinusitis EthmoidalisafiwahyuBelum ada peringkat
- AGRANULOSITOSISDokumen2 halamanAGRANULOSITOSISSarah AndrianiBelum ada peringkat
- PBL ImunologiDokumen25 halamanPBL ImunologiAndi Rizaldi Kurniawan MisbahBelum ada peringkat
- Tuberkulosis MilierDokumen7 halamanTuberkulosis MilierEva YunitaBelum ada peringkat
- Bone Turnover MarkersDokumen7 halamanBone Turnover MarkersEmmy HarefaBelum ada peringkat
- Referat Sjogren SyndromeDokumen40 halamanReferat Sjogren SyndromeLita BrilliandaBelum ada peringkat
- Tugas Responsi Mikro 2Dokumen8 halamanTugas Responsi Mikro 2KaroniBelum ada peringkat
- Judul TK PPDS Pulmo Update 1 Oktober 2019Dokumen53 halamanJudul TK PPDS Pulmo Update 1 Oktober 2019Aditya AlfariziBelum ada peringkat
- Imunologi: Skenario 2 Kelompok 11Dokumen22 halamanImunologi: Skenario 2 Kelompok 11rhizkyBelum ada peringkat
- Bagaimanakah Mekanisme Kerja Aminofillin Dalam Meningkatkan DiuresisDokumen1 halamanBagaimanakah Mekanisme Kerja Aminofillin Dalam Meningkatkan DiuresisTeresia MaharaniBelum ada peringkat
- Aslab ParasitologiDokumen3 halamanAslab ParasitologiRahma LuthfaBelum ada peringkat
- Laporan TutorialDokumen12 halamanLaporan Tutorialcar_sweetaBelum ada peringkat
- Emfisema AdalahDokumen3 halamanEmfisema AdalahBamz Hariyanto100% (4)
- PBL 18 - AsmaDokumen21 halamanPBL 18 - AsmaDefiita FiirdausBelum ada peringkat
- SkenarioDokumen2 halamanSkenarioFauzan AzhariBelum ada peringkat
- Sumber: Kamus Kedokteran Dorland, Edisi 29Dokumen24 halamanSumber: Kamus Kedokteran Dorland, Edisi 29tzz855Belum ada peringkat
- Wrap Up Skenario 3 Hemofilia - DichaDokumen24 halamanWrap Up Skenario 3 Hemofilia - DichaDicha Oseanni AndriswariBelum ada peringkat
- Laporan Kasus: Systemic Lupus Erytematous (SLE)Dokumen46 halamanLaporan Kasus: Systemic Lupus Erytematous (SLE)deladestianiaji2490Belum ada peringkat
- Laporan Referat VertigoDokumen17 halamanLaporan Referat VertigoMutiaraSartikaSuhardi100% (1)
- Rhinitis AlergiDokumen15 halamanRhinitis AlergiGilangBelum ada peringkat
- Interstitial Lung DiseaseDokumen24 halamanInterstitial Lung DiseaseShafira DianiBelum ada peringkat
- Tutorial 1Dokumen11 halamanTutorial 1Azmi SanjayaBelum ada peringkat
- Keracunan Alkohol AkutDokumen11 halamanKeracunan Alkohol AkutFauzan Luthfi A MBelum ada peringkat
- Radiologi RespirasiDokumen86 halamanRadiologi RespirasiintanrhamaBelum ada peringkat
- Laporan Kasus Depresi SedangDokumen22 halamanLaporan Kasus Depresi SedangFadyl Alannuari MBelum ada peringkat
- PATOGENESIS TUBERKULOSIS PARU DR JatuDokumen11 halamanPATOGENESIS TUBERKULOSIS PARU DR JatuAndy AShariBelum ada peringkat
- Ringkasan ImunDokumen20 halamanRingkasan ImunTimotius Ivan Sutanto0% (1)
- Respon Imun Tubuh Terhadap TuberkulosisDokumen6 halamanRespon Imun Tubuh Terhadap TuberkulosisBirgitta FajaraiBelum ada peringkat
- TB Dayu 1Dokumen13 halamanTB Dayu 1dayu reinaBelum ada peringkat
- Format Mikroplaning TUGAS1Dokumen45 halamanFormat Mikroplaning TUGAS1imatirpBelum ada peringkat
- Resume Modul NasionalismeDokumen8 halamanResume Modul NasionalismeimatirpBelum ada peringkat
- Wog 4.0Dokumen2 halamanWog 4.0imatirpBelum ada peringkat
- Tugas Manajemen LogistikDokumen1 halamanTugas Manajemen LogistikimatirpBelum ada peringkat
- Cover Cuci TanganDokumen1 halamanCover Cuci TanganimatirpBelum ada peringkat
- Tugas Manajemen LogistikDokumen1 halamanTugas Manajemen LogistikimatirpBelum ada peringkat
- Tugas Isu KontemporerDokumen5 halamanTugas Isu KontemporerimatirpBelum ada peringkat
- Resume Tugas Isu KontemporerDokumen9 halamanResume Tugas Isu KontemporerimatirpBelum ada peringkat
- Tugas EtikaDokumen3 halamanTugas EtikaimatirpBelum ada peringkat
- Tugas Etika Publik Kelompok 3Dokumen24 halamanTugas Etika Publik Kelompok 3imatirpBelum ada peringkat
- Tugas Etika Publik Kelompok 3Dokumen24 halamanTugas Etika Publik Kelompok 3imatirpBelum ada peringkat
- Etika OrganisasiDokumen10 halamanEtika OrganisasiimatirpBelum ada peringkat
- ETIKA BERNEGRA RezaDokumen3 halamanETIKA BERNEGRA RezaimatirpBelum ada peringkat
- ETIKA BERNEGRA RezaDokumen3 halamanETIKA BERNEGRA RezaimatirpBelum ada peringkat
- PenugasanDokumen1 halamanPenugasanimatirpBelum ada peringkat
- Resume Tugas Isu KontemporerDokumen9 halamanResume Tugas Isu KontemporerimatirpBelum ada peringkat
- Tugas Wasbang No.5Dokumen1 halamanTugas Wasbang No.5imatirpBelum ada peringkat
- Rizki Jadi EtikaDokumen9 halamanRizki Jadi EtikaimatirpBelum ada peringkat
- Tugas Wasbang No 4Dokumen1 halamanTugas Wasbang No 4imatirpBelum ada peringkat
- Kascil DR DhaniDokumen20 halamanKascil DR DhaniimatirpBelum ada peringkat
- PenugasanDokumen1 halamanPenugasanimatirpBelum ada peringkat
- Tugas Manajemen LogistikDokumen1 halamanTugas Manajemen LogistikimatirpBelum ada peringkat
- Presentasi Jurnal Magda-Tami DR Yulidar FIXDokumen22 halamanPresentasi Jurnal Magda-Tami DR Yulidar FIXimatirpBelum ada peringkat
- Kemenkes - Petunjuk Teknis Manajemen TB Anak (2013) PDFDokumen110 halamanKemenkes - Petunjuk Teknis Manajemen TB Anak (2013) PDFTita LuthfiaBelum ada peringkat
- Tugas Etika Diri SendiriDokumen2 halamanTugas Etika Diri SendiriimatirpBelum ada peringkat
- Pengobatan CMV TamiDokumen7 halamanPengobatan CMV TamiimatirpBelum ada peringkat
- SOAL UJIAN 3 CoasDokumen12 halamanSOAL UJIAN 3 CoasimatirpBelum ada peringkat
- ABSESDokumen5 halamanABSESDea Haykalsani HarahapBelum ada peringkat
- Lapkas Forensik AbortusDokumen30 halamanLapkas Forensik AbortusApul MunteBelum ada peringkat