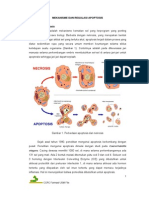
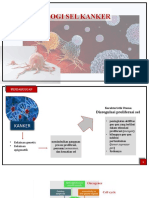
Anda mungkin juga menyukai
- Buku Masakan untuk Penyembuhan Skoliosis Anda: Jadikan tulang belakang lebih sehat dengan mengatur apa yang anda makan!Dari EverandBuku Masakan untuk Penyembuhan Skoliosis Anda: Jadikan tulang belakang lebih sehat dengan mengatur apa yang anda makan!Penilaian: 3.5 dari 5 bintang3.5/5 (4)
- APOPTOSISDokumen3 halamanAPOPTOSISAranty Fahira ArdisaBelum ada peringkat
- Biosel 5. ApoptosisDokumen27 halamanBiosel 5. ApoptosisilmisaBelum ada peringkat
- Mekanisme Dan Regulasi ApoptosisDokumen10 halamanMekanisme Dan Regulasi ApoptosisSusi DesmaryaniBelum ada peringkat
- Apaptosis Adalah Merupakan Kematian Sel Yang TerprogramDokumen5 halamanApaptosis Adalah Merupakan Kematian Sel Yang TerprogramAlimatul Misbah AlmunirohBelum ada peringkat
- Apoptosis Dan Proses PenuaanDokumen14 halamanApoptosis Dan Proses PenuaanAnieYuliyantiBelum ada peringkat
- Apoptosis MerupakanDokumen9 halamanApoptosis MerupakanDhifa CahyaniBelum ada peringkat
- Apoptosis Intrinsik, Intrinsic ApoptosisDokumen20 halamanApoptosis Intrinsik, Intrinsic Apoptosisdosendoktor100% (1)
- Definisi ApoptosisDokumen11 halamanDefinisi ApoptosisNuha RosyadaBelum ada peringkat
- ApoptosisDokumen24 halamanApoptosisFendy Hardian PermanaBelum ada peringkat
- Apoptosis Pada KankerDokumen2 halamanApoptosis Pada KankerDado ArmawanBelum ada peringkat
- Mekanisme Dan Regulasi Apoptosis1Dokumen17 halamanMekanisme Dan Regulasi Apoptosis1Aisah JuliantriBelum ada peringkat
- Apoptosis Dan NekrosisDokumen7 halamanApoptosis Dan NekrosisNaditya Fitriani HasanahBelum ada peringkat
- Laporan Apoptosis YulinarDokumen17 halamanLaporan Apoptosis YulinarYulinar Risky KaramanBelum ada peringkat
- Kematian SelDokumen9 halamanKematian SelAhmad HariyantoBelum ada peringkat
- Mekanisme Kematian SelDokumen6 halamanMekanisme Kematian SelDrgDondy100% (3)
- Jurnal ApoptosisDokumen6 halamanJurnal Apoptosispc komputerBelum ada peringkat
- Penuaan Dan Kematian SelDokumen47 halamanPenuaan Dan Kematian SelMi CheeBelum ada peringkat
- Kematian Sel Yang Terprogram PDFDokumen32 halamanKematian Sel Yang Terprogram PDFlisa fradisa100% (1)
- Kontrol Siklus SelDokumen22 halamanKontrol Siklus SelMuhammad Agus RizkiBelum ada peringkat
- 2Dokumen6 halaman2Feby FebryantiBelum ada peringkat
- Apoptosis Vs AutofagiDokumen15 halamanApoptosis Vs Autofagisaktiyonnie100% (3)
- Cedera Sel Dan Kematian Sel Dr. Komang Ardi Wahyuningsih, M.BiomedDokumen34 halamanCedera Sel Dan Kematian Sel Dr. Komang Ardi Wahyuningsih, M.BiomedPhilip PribadiBelum ada peringkat
- Apoptosis DirgaDokumen10 halamanApoptosis DirgaDirga AgungBelum ada peringkat
- ApoptosisDokumen15 halamanApoptosisAlyasa RamadhanBelum ada peringkat
- 1 ApoptosisDokumen35 halaman1 ApoptosisEvelyn AngieBelum ada peringkat
- 2013 Keseimbangan Apoptosis Dan Proliferasi SelDokumen13 halaman2013 Keseimbangan Apoptosis Dan Proliferasi Selkiki rawitriBelum ada peringkat
- Apoptopis PPTDokumen17 halamanApoptopis PPTSisy YuliantyBelum ada peringkat
- AntikankerDokumen41 halamanAntikankerIntanBelum ada peringkat
- ApoptosisDokumen48 halamanApoptosisAngga AriwijayaBelum ada peringkat
- Jurnal ReadingDokumen7 halamanJurnal ReadingMade paradiskaBelum ada peringkat
- Programmed Cell Death (APOPTOSIS)Dokumen26 halamanProgrammed Cell Death (APOPTOSIS)Rickky Kurniawan,MD100% (2)
- Cell Adaptation and Cell InjuryDokumen5 halamanCell Adaptation and Cell InjurySyachlaxxiBelum ada peringkat
- Referat Iskemik PenumbraDokumen13 halamanReferat Iskemik PenumbraWidya Widyarini0% (1)
- Program Kematian Sel (Apoptosis)Dokumen30 halamanProgram Kematian Sel (Apoptosis)Doni KurniawanBelum ada peringkat
- Tugas Farmakologi AntikankerDokumen42 halamanTugas Farmakologi AntikankerRohaniBelum ada peringkat
- Rohani - 1813105043 Antikanker PDFDokumen42 halamanRohani - 1813105043 Antikanker PDFRohaniBelum ada peringkat
- ApoptosisDokumen15 halamanApoptosisMuhammad HanifBelum ada peringkat
- Tugas Divisi Bedah Onkologi-KL (APOPTOSIS)Dokumen37 halamanTugas Divisi Bedah Onkologi-KL (APOPTOSIS)LafeniaBelum ada peringkat
- Biologi Sel ApoptosisDokumen15 halamanBiologi Sel ApoptosisYeni M. LianaBelum ada peringkat
- Biologi SelDokumen28 halamanBiologi SelEsa DesGaBelum ada peringkat
- Kuliah 14. ApoptosisDokumen20 halamanKuliah 14. Apoptosisratnakurniati_12Belum ada peringkat
- 4 Apoptosis and Cell DeathDokumen3 halaman4 Apoptosis and Cell DeathGina Kristina NanginBelum ada peringkat
- Apoptosis, Dr. AniDokumen42 halamanApoptosis, Dr. AniDebbie YourNitaBelum ada peringkat
- Radiobiologi Kelompok 3Dokumen12 halamanRadiobiologi Kelompok 3Rahmahh FyyBelum ada peringkat
- UntitledDokumen10 halamanUntitledRosalia MikasaBelum ada peringkat
- Apoptosis Dan NekrosisDokumen33 halamanApoptosis Dan NekrosisFiona BmfisBelum ada peringkat
- Artikel Biologi Sel Molekuler Tentang ApoptosisDokumen4 halamanArtikel Biologi Sel Molekuler Tentang ApoptosisDuta Nugraha Febrianto S1 - 2019Belum ada peringkat
- Biologi Sel KankerDokumen31 halamanBiologi Sel KankerTesa Yanuar RenjaniBelum ada peringkat
- ParafraseDokumen2 halamanParafraseroroBelum ada peringkat
- Apoptosis Adalah Suatu Proses Kematian Sel Yang TerprogramDokumen8 halamanApoptosis Adalah Suatu Proses Kematian Sel Yang TerprogramSean AndersonBelum ada peringkat
- Siklus Sel Dan KankerDokumen4 halamanSiklus Sel Dan KankerEka PujiastutiBelum ada peringkat
- Apoptosis FiixDokumen15 halamanApoptosis FiixDwi Ayni NahdaBelum ada peringkat
- Kematian Jaringan Dan Nekrosis Kelompok 4 Idk-1Dokumen34 halamanKematian Jaringan Dan Nekrosis Kelompok 4 Idk-1Chicilia Puspita DarmanigrumBelum ada peringkat
- Tugas Resume Pert.2 - Inayatul Maftukhah - 202205040 - Farmasi 1B - Perbedaan Nekrosis & ApoptosisDokumen4 halamanTugas Resume Pert.2 - Inayatul Maftukhah - 202205040 - Farmasi 1B - Perbedaan Nekrosis & ApoptosisInayatul MaftukhahBelum ada peringkat
- Jalur MitokondriaDokumen3 halamanJalur MitokondriaDwi Amalia husnaBelum ada peringkat
- Ihsan Febrianto Rahman - B4 - Makalah Tutorial Case 1 (FBS 2) Jejas Sel, Adaptasi Sel, Reaksi Radang, Dan Pemulihan JaringanDokumen64 halamanIhsan Febrianto Rahman - B4 - Makalah Tutorial Case 1 (FBS 2) Jejas Sel, Adaptasi Sel, Reaksi Radang, Dan Pemulihan JaringanIhsan KBelum ada peringkat
- ApoptosisDokumen31 halamanApoptosisAhmad Faizal BustomiBelum ada peringkat
- CUB Makalah Donor SafetyDokumen3 halamanCUB Makalah Donor SafetyTeofilus TrichinasBelum ada peringkat
- JesicaLubertina - PPT KomplemenDokumen8 halamanJesicaLubertina - PPT KomplemenTeofilus TrichinasBelum ada peringkat
- Jesica Lubertina - 211206011 - Link Foto Dan VideoDokumen1 halamanJesica Lubertina - 211206011 - Link Foto Dan VideoTeofilus TrichinasBelum ada peringkat
- Jesica Lubertina - 211206011 - Kuis Teori SGD Pertemuan 2Dokumen5 halamanJesica Lubertina - 211206011 - Kuis Teori SGD Pertemuan 2Teofilus TrichinasBelum ada peringkat
- Tugas Praktikum 2 CUB 2022Dokumen1 halamanTugas Praktikum 2 CUB 2022Teofilus TrichinasBelum ada peringkat
- TBD - Judul Dan Rumusan Masalah Sebelum RevisiDokumen4 halamanTBD - Judul Dan Rumusan Masalah Sebelum RevisiTeofilus TrichinasBelum ada peringkat
- JesicaLubertina - 211206011 - Tugas Praktik Temu 2 Kegiatan 1 Bahasa IndonesiaDokumen3 halamanJesicaLubertina - 211206011 - Tugas Praktik Temu 2 Kegiatan 1 Bahasa IndonesiaTeofilus TrichinasBelum ada peringkat
- Psikologi KomunikasiDokumen306 halamanPsikologi KomunikasiALWIN NUGRAHA100% (1)
- JesicaLubertina - 211206011 - Argumentatif Donor DarahDokumen3 halamanJesicaLubertina - 211206011 - Argumentatif Donor DarahTeofilus TrichinasBelum ada peringkat
- JesicaLubertina 211206011 AnekdotDokumen2 halamanJesicaLubertina 211206011 AnekdotTeofilus TrichinasBelum ada peringkat
- Psikologi KomunikasiDokumen306 halamanPsikologi KomunikasiALWIN NUGRAHA100% (1)
- Pelaksanaan Pancasila Sebagai Pedoman Pengembangan IptekDokumen6 halamanPelaksanaan Pancasila Sebagai Pedoman Pengembangan IptekTeofilus TrichinasBelum ada peringkat
- Modul Psikologi KomunikasiDokumen41 halamanModul Psikologi Komunikasidevy oktavianiBelum ada peringkat
- Modul Psikologi KomunikasiDokumen41 halamanModul Psikologi Komunikasidevy oktavianiBelum ada peringkat
- Pelaksanaan Pancasila Sebagai Pedoman Pengembangan IptekDokumen6 halamanPelaksanaan Pancasila Sebagai Pedoman Pengembangan IptekTeofilus TrichinasBelum ada peringkat
- Tugas IMUNO 02 - Sistem Komplemen Dan JalurnyaDokumen7 halamanTugas IMUNO 02 - Sistem Komplemen Dan JalurnyaJessica CaeciliaBelum ada peringkat
- Kajian Ilmiah Issue Nasional FixDokumen7 halamanKajian Ilmiah Issue Nasional FixBella datunsolangBelum ada peringkat
- FibrinolisisDokumen5 halamanFibrinolisisTeofilus TrichinasBelum ada peringkat
- Denah LaboratoriumDokumen3 halamanDenah LaboratoriumTeofilus TrichinasBelum ada peringkat
- Proses Pemekuan Darah Dan Fibrinolitik 8 November 2021Dokumen41 halamanProses Pemekuan Darah Dan Fibrinolitik 8 November 2021Teofilus TrichinasBelum ada peringkat
- Soal Kredensial LaboratoriumDokumen7 halamanSoal Kredensial LaboratoriumTeofilus TrichinasBelum ada peringkat
- 501 26 PB PDFDokumen8 halaman501 26 PB PDFsilviBelum ada peringkat
- Chapter IIDokumen19 halamanChapter IIIzfaningrum Melati SukmaBelum ada peringkat
- Materi Kuliah Ida Bagus Suardana 1503977453 PDFDokumen36 halamanMateri Kuliah Ida Bagus Suardana 1503977453 PDFWilyams TutuhatunewaBelum ada peringkat
- Limporetikuler - MK ImunologiDokumen19 halamanLimporetikuler - MK ImunologiTeofilus TrichinasBelum ada peringkat
- Sistem Limfoid Primer Dan Sekunder - NewDokumen10 halamanSistem Limfoid Primer Dan Sekunder - NewLab bethamedikaBelum ada peringkat
- Makalah Siklus Sel PDFDokumen34 halamanMakalah Siklus Sel PDFYulinar Risky KaramanBelum ada peringkat
- Makalah BioselDokumen22 halamanMakalah BioselAchmad RodiansyahBelum ada peringkat
- Praktikum LimporetikulerDokumen1 halamanPraktikum LimporetikulerTeofilus TrichinasBelum ada peringkat
- Regulasi Siklus SelDokumen35 halamanRegulasi Siklus SelTeofilus TrichinasBelum ada peringkat