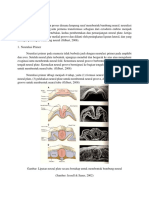
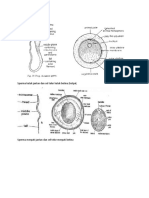
Anda mungkin juga menyukai
- Asimilasi NitrogenDokumen4 halamanAsimilasi Nitrogenrina putriBelum ada peringkat
- Daur NitrogenDokumen3 halamanDaur NitrogenGrecencia SiraitBelum ada peringkat
- Makalah Kelompok 8 Fiksasi Dan Asimilasi NitrogenDokumen16 halamanMakalah Kelompok 8 Fiksasi Dan Asimilasi NitrogenMaria Kordak100% (2)
- Daur NitrogenDokumen4 halamanDaur NitrogenRizkyahBelum ada peringkat
- Daur NitrogenDokumen4 halamanDaur NitrogenRIZKYAH PUTRI AMALIA mhsD3KL2019BBelum ada peringkat
- Makalah Siklus MikrobaDokumen11 halamanMakalah Siklus Mikrobasamrotul puadahBelum ada peringkat
- Siklus Nitrogen Dengan Mengaitkan Keberadaan Nitrogen Di Udara, Tanah, Dan Makhluk HidupDokumen3 halamanSiklus Nitrogen Dengan Mengaitkan Keberadaan Nitrogen Di Udara, Tanah, Dan Makhluk Hiduprita tri maharaniBelum ada peringkat
- Nitrogen 2Dokumen31 halamanNitrogen 2Muhammad Hani Azhary0% (1)
- KELOMPOK 1-PPT SIKLUS NITROGEN DALAM TANAH DAN REDUKSI NITRAT-MIKROBIOLOGI TANAH-dikonversiDokumen13 halamanKELOMPOK 1-PPT SIKLUS NITROGEN DALAM TANAH DAN REDUKSI NITRAT-MIKROBIOLOGI TANAH-dikonversiResti YuliaBelum ada peringkat
- Metabolisme NitrogenDokumen13 halamanMetabolisme NitrogenAnonymous a3btz4Belum ada peringkat
- Daur NitrogenDokumen2 halamanDaur NitrogenmqkomputerBelum ada peringkat
- Tgs 1Dokumen8 halamanTgs 1Tamauli NababanBelum ada peringkat
- Nutrisi Tanaman M.jais 20011014052Dokumen4 halamanNutrisi Tanaman M.jais 20011014052Muhammad JaisBelum ada peringkat
- Daur BiokimiaDokumen17 halamanDaur BiokimiaRetno NusantariBelum ada peringkat
- Daur FosforDokumen10 halamanDaur FosforReeza MegavitryBelum ada peringkat
- Makalah Fistum KLP 6Dokumen16 halamanMakalah Fistum KLP 6Marisa RintaniaBelum ada peringkat
- Daur Nitroge1Dokumen5 halamanDaur Nitroge1Muhammad Faris Rasyad SiddiqBelum ada peringkat
- Fiksasi NDokumen10 halamanFiksasi NGita MaizaBelum ada peringkat
- Makalah Biokimia TanamanDokumen9 halamanMakalah Biokimia TanamanCandra Irawan100% (1)
- Dea Gita 431419062 Rangkuman Siklus NitrogenDokumen7 halamanDea Gita 431419062 Rangkuman Siklus NitrogenDea GitaBelum ada peringkat
- Daur Nitrogen Samsons Xa1Dokumen2 halamanDaur Nitrogen Samsons Xa1samsonsimbolonBelum ada peringkat
- Bakteri NitrifikasiDokumen8 halamanBakteri NitrifikasirikajuliskaBelum ada peringkat
- Pembahasan MEtabolisme NitrogenDokumen12 halamanPembahasan MEtabolisme NitrogenEireine Injiia LiowBelum ada peringkat
- Siklus Karbon:: 1. Proses FotosintesisDokumen4 halamanSiklus Karbon:: 1. Proses FotosintesisRaghil MahardikaBelum ada peringkat
- NITROGENDokumen7 halamanNITROGENYoga MuftiBelum ada peringkat
- HumusDokumen6 halamanHumusAbdul AzizBelum ada peringkat
- HumusDokumen6 halamanHumusAbdul AzizBelum ada peringkat
- Daur NitrogenDokumen3 halamanDaur NitrogenYudha Maulidza Al FariziBelum ada peringkat
- Tugas Klipping 1 Kimia LingkunganDokumen14 halamanTugas Klipping 1 Kimia LingkunganFauzia YunsyahanaBelum ada peringkat
- AsimilasiDokumen5 halamanAsimilasiprantottiBelum ada peringkat
- 7 Mikrobiologi Lingkungan-RevDokumen38 halaman7 Mikrobiologi Lingkungan-RevHendra RonaldiBelum ada peringkat
- Siklus Dan FiksasiDokumen27 halamanSiklus Dan FiksasiSyarif Hidayat AmrullahBelum ada peringkat
- Asimilasi Nitrogen Dan BelerangDokumen12 halamanAsimilasi Nitrogen Dan BelerangReksi Bayu El Hanif0% (1)
- Daur NitrogenDokumen3 halamanDaur NitrogenAfifahBelum ada peringkat
- D - Kelompok 3 - PPT Daur NitrogenDokumen14 halamanD - Kelompok 3 - PPT Daur NitrogenNisa ChaeraBelum ada peringkat
- Paper Peranan Mikroba Dalam Transformasi NitrogenDokumen11 halamanPaper Peranan Mikroba Dalam Transformasi NitrogenDessy Djata NduaBelum ada peringkat
- DaurDokumen8 halamanDaurAnggia RahayuBelum ada peringkat
- Daur Nitrogen AyyuDokumen9 halamanDaur Nitrogen AyyuAyu AnggrainiBelum ada peringkat
- Penjelasan Proses Siklus Nitrogen Dalam EkosistemDokumen5 halamanPenjelasan Proses Siklus Nitrogen Dalam EkosistemAdilhaJhaBelum ada peringkat
- Metabolisme SulfurDokumen19 halamanMetabolisme SulfurAndreas Da Silva100% (2)
- Asimilasi Nitrogen Dan SulfurDokumen11 halamanAsimilasi Nitrogen Dan SulfurBadrus Al FaìruzBelum ada peringkat
- Daur BiogeokimiaDokumen9 halamanDaur BiogeokimiaAnnis MulyaniBelum ada peringkat
- DAUR NitrogenDokumen6 halamanDAUR NitrogenMuhammad Hanif RasydaBelum ada peringkat
- Tugas Fiksasi NDokumen25 halamanTugas Fiksasi NAndiAlfianiBelum ada peringkat
- Siklus NitrogenDokumen8 halamanSiklus NitrogenTriRahmadhaniBelum ada peringkat
- Metabolisme NitrogenDokumen14 halamanMetabolisme NitrogenFitri Utami Hasan100% (3)
- Siklus NitrogenDokumen7 halamanSiklus NitrogenAnang BudiBelum ada peringkat
- PresentasiDokumen9 halamanPresentasiYossi CuyBelum ada peringkat
- Mekanisme Penambatan Nitrogen Oleh Bakteri Bintil AkarDokumen4 halamanMekanisme Penambatan Nitrogen Oleh Bakteri Bintil Akarayu lestariBelum ada peringkat
- Siklus BiogeokimiaDokumen6 halamanSiklus Biogeokimiacrissa estellaBelum ada peringkat
- Tugas BiogeokimiaDokumen9 halamanTugas BiogeokimiaMiskah OberonBelum ada peringkat
- Siklus NitrogenDokumen2 halamanSiklus Nitrogen14Fira Sofia Ratu AfandiBelum ada peringkat
- BiogeokimiaDokumen20 halamanBiogeokimiaDewa Ayu Mery AgustinBelum ada peringkat
- Pengertian Daur NitrogenDokumen3 halamanPengertian Daur NitrogenZaid ZaidBelum ada peringkat
- Nitrogen - Nur Septiana GobelDokumen3 halamanNitrogen - Nur Septiana GobelTiyaBelum ada peringkat
- Mikrodas (Belum Siap)Dokumen2 halamanMikrodas (Belum Siap)M. Dandi Arsio Zikri 2104124862Belum ada peringkat
- Tanaman Pepohonan Untuk Menjernihkan & Menetralisir Air Limbah Beracun Berbahaya Dari Kawasan Perairan Laut Sungai DanauDari EverandTanaman Pepohonan Untuk Menjernihkan & Menetralisir Air Limbah Beracun Berbahaya Dari Kawasan Perairan Laut Sungai DanauBelum ada peringkat
- Tri TerpenoidDokumen5 halamanTri Terpenoidida nurpitaBelum ada peringkat
- Halaman I II III IV, Tinggal Ngasih HalamanDokumen3 halamanHalaman I II III IV, Tinggal Ngasih Halamanida nurpitaBelum ada peringkat
- Skrining FitokimiaDokumen1 halamanSkrining Fitokimiaida nurpitaBelum ada peringkat
- Metode Skrining Fitokimia Digunakan Untuk Mengetahui Kandungan Metabolit SekunderDokumen1 halamanMetode Skrining Fitokimia Digunakan Untuk Mengetahui Kandungan Metabolit Sekunderida nurpitaBelum ada peringkat
- Diversitas Antibod1 1Dokumen3 halamanDiversitas Antibod1 1ida nurpitaBelum ada peringkat
- Resume Istilah ParasitologiDokumen20 halamanResume Istilah Parasitologiida nurpitaBelum ada peringkat
- KH Tabel PerbandinganDokumen15 halamanKH Tabel Perbandinganida nurpita0% (1)
- Rujukan KH AvesDokumen8 halamanRujukan KH Avesida nurpitaBelum ada peringkat
- Metode Skrining Fitokimia Digunakan Untuk Mengetahui Kandungan Metabolit SekunderDokumen1 halamanMetode Skrining Fitokimia Digunakan Untuk Mengetahui Kandungan Metabolit Sekunderida nurpitaBelum ada peringkat
- Pembahasan PHDokumen7 halamanPembahasan PHida nurpitaBelum ada peringkat
- Factsheet WWF Indonesia Rangkong Badak PDFDokumen2 halamanFactsheet WWF Indonesia Rangkong Badak PDFida nurpitaBelum ada peringkat
- Asteroidea OphiuroideaDokumen7 halamanAsteroidea Ophiuroideaida nurpitaBelum ada peringkat
- Kata PengantarDokumen2 halamanKata Pengantarida nurpitaBelum ada peringkat
- PudjiadiDokumen1 halamanPudjiadiida nurpitaBelum ada peringkat
- Poin 2Dokumen5 halamanPoin 2ida nurpitaBelum ada peringkat
- Kelompok Ikan - Kelompok 1Dokumen58 halamanKelompok Ikan - Kelompok 1ida nurpitaBelum ada peringkat
- Daftar Rujukan Tekanan Jantung Dan VenaDokumen5 halamanDaftar Rujukan Tekanan Jantung Dan Venaida nurpitaBelum ada peringkat
- HolothuroideaDokumen2 halamanHolothuroideaida nurpitaBelum ada peringkat
- Peranan MikroDokumen4 halamanPeranan Mikroida nurpitaBelum ada peringkat
- NeurulasiDokumen5 halamanNeurulasiida nurpitaBelum ada peringkat
- 4Dokumen4 halaman4ida nurpitaBelum ada peringkat
- Sperma Katak Jantan Dan Sel Telur Katak BetinaDokumen2 halamanSperma Katak Jantan Dan Sel Telur Katak Betinaida nurpitaBelum ada peringkat
- Daftar PustakaDokumen3 halamanDaftar Pustakaida nurpitaBelum ada peringkat
- Resume Materi Griffith DKKDokumen3 halamanResume Materi Griffith DKKida nurpitaBelum ada peringkat
- Data Pith FallDokumen8 halamanData Pith Fallida nurpitaBelum ada peringkat
- Pembahasan Gerak BakteriDokumen2 halamanPembahasan Gerak Bakteriida nurpitaBelum ada peringkat
- Diskusi, Kesipulan, Dapus ALTDokumen1 halamanDiskusi, Kesipulan, Dapus ALTida nurpitaBelum ada peringkat
- Operonlakpdf PDFDokumen5 halamanOperonlakpdf PDFida nurpitaBelum ada peringkat
- Peranan MikroDokumen4 halamanPeranan Mikroida nurpitaBelum ada peringkat
- Tulang KerasDokumen16 halamanTulang Kerasida nurpitaBelum ada peringkat