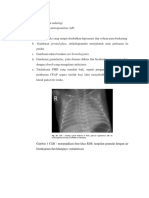
Anda mungkin juga menyukai
- Sistem Imun KomplemenDokumen28 halamanSistem Imun KomplemenMarLeniRNBelum ada peringkat
- SISTEM KOMPLEMENDokumen8 halamanSISTEM KOMPLEMENilhamhamka12345Belum ada peringkat
- Manfaat AsiDokumen6 halamanManfaat AsighearaudhaBelum ada peringkat
- Infeksi HookwormDokumen5 halamanInfeksi HookwormDoctorKawaii100% (1)
- HemostasisDokumen64 halamanHemostasisSry Rahayu100% (1)
- LIMFOMA-ANAKDokumen29 halamanLIMFOMA-ANAKNurulBelum ada peringkat
- ARAS dan Penurunan KesadaranDokumen15 halamanARAS dan Penurunan KesadaranmamanBelum ada peringkat
- HematopoiesisDokumen4 halamanHematopoiesislisna_fk09Belum ada peringkat
- Presentation SifilisDokumen14 halamanPresentation SifilisrullynmandarBelum ada peringkat
- Kelompok 4 (SATPHYLOCOCCUS EPIDEERMIDIS)Dokumen6 halamanKelompok 4 (SATPHYLOCOCCUS EPIDEERMIDIS)Bang SanssBelum ada peringkat
- TOF AnakDokumen11 halamanTOF Anaksang bangauBelum ada peringkat
- kELOMPOK 4 - APLIKASI PRAKTIS IMUNOLOGIDokumen77 halamankELOMPOK 4 - APLIKASI PRAKTIS IMUNOLOGITiara MuzadilahBelum ada peringkat
- Definisi Fluor AlbusDokumen10 halamanDefinisi Fluor AlbusAnonymous VwqvACBelum ada peringkat
- Bab I KandidiasisDokumen14 halamanBab I KandidiasisIrma SetyawatiBelum ada peringkat
- IMUNOLOGIDokumen2 halamanIMUNOLOGIMiftakhur RohmahBelum ada peringkat
- Eritropoiesis dalam Sumsum TulangDokumen6 halamanEritropoiesis dalam Sumsum TulangRagilBelum ada peringkat
- Mikrobiota Saluran CernaDokumen5 halamanMikrobiota Saluran CernaNailiBelum ada peringkat
- Bab IiDokumen17 halamanBab IiRully SyahrizalBelum ada peringkat
- Disseminated Intravascular Coagulation (DIC)Dokumen23 halamanDisseminated Intravascular Coagulation (DIC)YuniBelum ada peringkat
- HEMOPOIESISDokumen7 halamanHEMOPOIESISJean StepaniBelum ada peringkat
- Fisiologi Lymph NodeDokumen2 halamanFisiologi Lymph NodeLismi Awliya RahmahBelum ada peringkat
- Laporan Tutorial Kelompok 5 Skenario 3 Blok 5Dokumen32 halamanLaporan Tutorial Kelompok 5 Skenario 3 Blok 5Angga negaraBelum ada peringkat
- Ringkasan CESTODADokumen16 halamanRingkasan CESTODARizqon Yassir KuswondoBelum ada peringkat
- ImunoterapiDokumen6 halamanImunoterapiarmasriBelum ada peringkat
- ANEMIA HEMOLITIK: PENGERTIAN, PENYEBAB, GEJALA, DAN PENANGANANNYADokumen15 halamanANEMIA HEMOLITIK: PENGERTIAN, PENYEBAB, GEJALA, DAN PENANGANANNYAAndy AbadiBelum ada peringkat
- IMUNODEFISIENSIDokumen13 halamanIMUNODEFISIENSIwinda mulyaBelum ada peringkat
- Sel Natural KillerDokumen2 halamanSel Natural KillerRio Fernandes Tan100% (1)
- Sitokin & KemokinDokumen15 halamanSitokin & KemokinAhmat Rediansya PutraBelum ada peringkat
- Program Kematian Sel (Apoptosis)Dokumen30 halamanProgram Kematian Sel (Apoptosis)Doni KurniawanBelum ada peringkat
- Penyakit Graves: Penyebab Hipertiroid Paling SeringDokumen9 halamanPenyakit Graves: Penyebab Hipertiroid Paling SeringGusti Ngurah PBelum ada peringkat
- Sistem Imun KulitDokumen35 halamanSistem Imun KulitNew Fiza Faizatul IzzaBelum ada peringkat
- Faktor KOAGULASIDokumen4 halamanFaktor KOAGULASIMelany Amdira IIBelum ada peringkat
- Histologi DarahDokumen7 halamanHistologi DarahIndah Dewi OktaviaBelum ada peringkat
- TGF BetaDokumen14 halamanTGF BetaMuhammad RizalBelum ada peringkat
- Aterosklerosis Merupakan Proses Pembentukan PlakDokumen3 halamanAterosklerosis Merupakan Proses Pembentukan PlakNovera WidiastutyBelum ada peringkat
- Limpa/lienDokumen3 halamanLimpa/lientasyadelizaBelum ada peringkat
- GranulopoesisilDokumen9 halamanGranulopoesisilyuniBelum ada peringkat
- Anatomi Histologi Fisiologi MammaeDokumen18 halamanAnatomi Histologi Fisiologi MammaeAdidya Rizky PambudiBelum ada peringkat
- Sussy - Metabolisme Asam FolatDokumen15 halamanSussy - Metabolisme Asam FolatAlkanio AlfanioBelum ada peringkat
- KELAINAN KROMOSOMDokumen9 halamanKELAINAN KROMOSOMlimBelum ada peringkat
- 2a - Anugerah Bataha - Hematologi 2-Ringkasan Fragilitas Eritrosit PDFDokumen3 halaman2a - Anugerah Bataha - Hematologi 2-Ringkasan Fragilitas Eritrosit PDFCherulo SombaBelum ada peringkat
- Acute Myeloid LeukemiaDokumen10 halamanAcute Myeloid Leukemiailham pratamaBelum ada peringkat
- Trichomonas VaginalisDokumen6 halamanTrichomonas VaginalisResmha AndhikaBelum ada peringkat
- Tumbuh Kembang RemajaDokumen104 halamanTumbuh Kembang RemajaShodiqul AminBelum ada peringkat
- Laporan Tutorial Modul 4 PerdarahanDokumen58 halamanLaporan Tutorial Modul 4 PerdarahanAulia Kusumawati MerimbanoBelum ada peringkat
- Makalah Autoimunitas Atika Jaya Rani (13330716)Dokumen46 halamanMakalah Autoimunitas Atika Jaya Rani (13330716)SwerysBelum ada peringkat
- Trichomoniasis Vaginalis 2012Dokumen24 halamanTrichomoniasis Vaginalis 2012Eka Yusi Nur Athiyyah100% (1)
- Virus Zika On GoingDokumen16 halamanVirus Zika On GoingPurnomo Ponco NugrohoBelum ada peringkat
- GlikokaliksDokumen1 halamanGlikokaliksFatur RahmnBelum ada peringkat
- BBDM Histologi Genitalia WanitaDokumen26 halamanBBDM Histologi Genitalia WanitaaidillahBelum ada peringkat
- Viii. Proses Invasi Dan Metastasis Tumor GanasDokumen3 halamanViii. Proses Invasi Dan Metastasis Tumor GanasghearaudhaBelum ada peringkat
- Plasenta PreviaDokumen12 halamanPlasenta PreviaResi Lystianto PutraBelum ada peringkat
- NLRDokumen5 halamanNLRarviantyintan100% (1)
- Hemosistein dan Penyakit JantungDokumen12 halamanHemosistein dan Penyakit JantungEka Hardiyanti HusainBelum ada peringkat
- Laporan Diskusi Kelompok P3 Kelompok 1 Hematologi Onkologi PDFDokumen49 halamanLaporan Diskusi Kelompok P3 Kelompok 1 Hematologi Onkologi PDFNavisaBelum ada peringkat
- HEMATOPOIESIS PROSESDokumen6 halamanHEMATOPOIESIS PROSESRasmaah MukhtarBelum ada peringkat
- Pemeriksaan Radiologi HMDDokumen6 halamanPemeriksaan Radiologi HMDanggrenii SBelum ada peringkat
- GIGANTISMEDokumen39 halamanGIGANTISMERicoPratamaWiyonoBelum ada peringkat
- Imunologi KomplemenDokumen26 halamanImunologi KomplemenAyu RukminiBelum ada peringkat
- Kuliah 2 - Sistem Komplemen, Sitokin Dan Imunitas Humoral-Selular - MBKMDokumen82 halamanKuliah 2 - Sistem Komplemen, Sitokin Dan Imunitas Humoral-Selular - MBKMREGITA RAHMA FISKABelum ada peringkat
- 6316 18779 1 PBDokumen16 halaman6316 18779 1 PBRizkyBelum ada peringkat
- Pembimbingan Tugas Akhir BiologiDokumen3 halamanPembimbingan Tugas Akhir BiologiRizkyBelum ada peringkat
- 7004 20913 1 PBDokumen6 halaman7004 20913 1 PBRizkyBelum ada peringkat
- Surat LemburDokumen1 halamanSurat LemburRizkyBelum ada peringkat
- Efek Antidiabetes Kombinasi Ekstrak Pandan dan SalamDokumen17 halamanEfek Antidiabetes Kombinasi Ekstrak Pandan dan SalamRizkyBelum ada peringkat
- TSV PADA UDANGDokumen2 halamanTSV PADA UDANGRizkyBelum ada peringkat
- 02 Peraturan ItpDokumen2 halaman02 Peraturan ItpRizkyBelum ada peringkat
- Dian Fajar Ariyani M0407030Dokumen67 halamanDian Fajar Ariyani M0407030RizkyBelum ada peringkat
- Makalah Pencling Pencemaran UdaraDokumen14 halamanMakalah Pencling Pencemaran UdaraRizkyBelum ada peringkat
- Kajian Morfologi Dan Review Fitokimia Tumbuhan Kersen (Muntingia Calabura L)Dokumen7 halamanKajian Morfologi Dan Review Fitokimia Tumbuhan Kersen (Muntingia Calabura L)RizkyBelum ada peringkat
- Surat PenundaanDokumen2 halamanSurat PenundaanRizkyBelum ada peringkat
- Lampiran Dokumentasi Purifikasi DNA Shift BDokumen2 halamanLampiran Dokumentasi Purifikasi DNA Shift BRizkyBelum ada peringkat
- JUDULDokumen26 halamanJUDULRizkyBelum ada peringkat
- BVT GejalaDokumen2 halamanBVT GejalaRizkyBelum ada peringkat
- TSV PADA UDANGDokumen2 halamanTSV PADA UDANGRizkyBelum ada peringkat
- Laporan KMM Buat PsDokumen31 halamanLaporan KMM Buat PsRizkyBelum ada peringkat
- Revisi Laporan MagangDokumen30 halamanRevisi Laporan MagangRizkyBelum ada peringkat
- Punya BagusDokumen2 halamanPunya BagusRizkyBelum ada peringkat
- ISOLASI BAKTERIDokumen31 halamanISOLASI BAKTERIRizkyBelum ada peringkat
- Ekosistem NokturnalDokumen5 halamanEkosistem NokturnalRizky100% (1)
- Acara 4Dokumen3 halamanAcara 4RizkyBelum ada peringkat
- Daur KarbonDokumen4 halamanDaur KarbonRizkyBelum ada peringkat
- BioterorismeDokumen16 halamanBioterorismeRizkyBelum ada peringkat
- OUTLINEDokumen6 halamanOUTLINERizkyBelum ada peringkat
- UTS Kesling 2017Dokumen1 halamanUTS Kesling 2017RizkyBelum ada peringkat
- Air Beras-1Dokumen13 halamanAir Beras-1RizkyBelum ada peringkat
- Air Beras-1Dokumen13 halamanAir Beras-1RizkyBelum ada peringkat
- KolesistokininDokumen2 halamanKolesistokininRizky100% (1)
- Air Beras-1Dokumen13 halamanAir Beras-1RizkyBelum ada peringkat