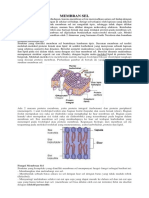
Anda mungkin juga menyukai
- Tugas KLP 4-Struktur & Mekanisme Transfor Melalui MembranDokumen23 halamanTugas KLP 4-Struktur & Mekanisme Transfor Melalui MembranYuri GagarinBelum ada peringkat
- Anatomi - Membran PlasmaDokumen11 halamanAnatomi - Membran PlasmaMia100% (1)
- Membran SelDokumen7 halamanMembran SelMirazizaAmandaBelum ada peringkat
- Struktur Lipid Bilayer Membran SelDokumen11 halamanStruktur Lipid Bilayer Membran Selshofa_aditya100% (1)
- Makalah Biomed Kelompok 3Dokumen12 halamanMakalah Biomed Kelompok 3Ayu SivaBelum ada peringkat
- Tugas IdkDokumen45 halamanTugas Idkana khulil jannahBelum ada peringkat
- Membran PlasmaDokumen19 halamanMembran PlasmaAndhika Perkasa SumartoBelum ada peringkat
- Biosel Kelompok 3 (Revisi)Dokumen33 halamanBiosel Kelompok 3 (Revisi)Annisa RachmieBelum ada peringkat
- MEMBRANDokumen35 halamanMEMBRANMuhammad PutraBelum ada peringkat
- Membran Sel StrukturDokumen6 halamanMembran Sel StrukturNurfaiziBelum ada peringkat
- Membran Sel 1Dokumen5 halamanMembran Sel 1Ibana Rabbiatul AmarinaBelum ada peringkat
- Resume Tentang Membran Plasma SelDokumen17 halamanResume Tentang Membran Plasma SelSely BoysalaBelum ada peringkat
- Membran Sel Dan Transportasi SelDokumen11 halamanMembran Sel Dan Transportasi SelIndri HardiyantiBelum ada peringkat
- Membran SelDokumen27 halamanMembran SelMuhammad Budi SyahputraBelum ada peringkat
- Materi PPT Struktur TumbuhanDokumen21 halamanMateri PPT Struktur TumbuhanDunia KarirBelum ada peringkat
- Paper Organel Sel (Agus P)Dokumen32 halamanPaper Organel Sel (Agus P)DuvanAgaveSnrBelum ada peringkat
- Makalah Struktur Umum Membran SelDokumen14 halamanMakalah Struktur Umum Membran SelReynanda ApriliaBelum ada peringkat
- Membran PlasmaDokumen8 halamanMembran PlasmaAndhika Perkasa SumartoBelum ada peringkat
- Membran Sel: Struktur, Komposisi, dan FungsinyaDokumen8 halamanMembran Sel: Struktur, Komposisi, dan FungsinyaGanendra Awang KristandyaBelum ada peringkat
- Bu ArniDokumen41 halamanBu Arnititin dewi sartika silabanBelum ada peringkat
- Teori MozaikDokumen9 halamanTeori Mozaikasyrafun nisaBelum ada peringkat
- Makalah Membran PlasmaDokumen26 halamanMakalah Membran PlasmaLanna Murpi Pertiwi0% (1)
- Biosel FixDokumen13 halamanBiosel Fixwulan rahayuBelum ada peringkat
- LKS 5 NonDokumen8 halamanLKS 5 NongissaBelum ada peringkat
- Pengertian Membran SelDokumen6 halamanPengertian Membran SelRut NguruBelum ada peringkat
- Makalah Gabungan Membran SelDokumen67 halamanMakalah Gabungan Membran SelHaryatin Nurul AfifahBelum ada peringkat
- MEMBRAN STRUKTUR & FUNGSIDokumen28 halamanMEMBRAN STRUKTUR & FUNGSIkukun0% (1)
- Membran SelDokumen15 halamanMembran SelRisnawatiHRBelum ada peringkat
- MEMBRAN SELDokumen20 halamanMEMBRAN SELNurul Mukhlisa AmirBelum ada peringkat
- Struktur Membran SelDokumen5 halamanStruktur Membran Selmariam_n10tangsel100% (1)
- Struktur Dan Fungsi MembranDokumen12 halamanStruktur Dan Fungsi MembranNurul MentariBelum ada peringkat
- Makalah Membran PlasmaDokumen8 halamanMakalah Membran PlasmaYesi FitrianiBelum ada peringkat
- MEMBRAN SELDokumen30 halamanMEMBRAN SELK-NndafBelum ada peringkat
- Transport Membran SelDokumen15 halamanTransport Membran SelMaria PreciliaBelum ada peringkat
- MEMBRAN FUNGSIDokumen16 halamanMEMBRAN FUNGSIindicha0905Belum ada peringkat
- PeroksisomDokumen21 halamanPeroksisomRahmaAswagBelum ada peringkat
- Struktur-Sel 2Dokumen12 halamanStruktur-Sel 2IswerBelum ada peringkat
- Struktur Dan Fungsi Membran SelDokumen16 halamanStruktur Dan Fungsi Membran SelNURWIDAYANTI100% (3)
- Struktur Dan Fungsi Membran SelDokumen10 halamanStruktur Dan Fungsi Membran SelKejora HanadinantiBelum ada peringkat
- Makalah 2 Membran SelDokumen28 halamanMakalah 2 Membran SelMurditya Kemal VasyaBelum ada peringkat
- Struktur Dan Fungsi Membran SelDokumen7 halamanStruktur Dan Fungsi Membran SelCintaBelum ada peringkat
- KJ010 4 7407 FBM111 092018 DocDokumen16 halamanKJ010 4 7407 FBM111 092018 DocNabila Azzah MahdiyahBelum ada peringkat
- Pengambilan Nutrisi Dan AsimilasiDokumen8 halamanPengambilan Nutrisi Dan AsimilasijenisatrianiBelum ada peringkat
- Struktur Komponen Membran SelDokumen7 halamanStruktur Komponen Membran SelWahyu HidayatBelum ada peringkat
- Albert 617-619Dokumen4 halamanAlbert 617-619arif hidayatBelum ada peringkat
- Membran SelDokumen40 halamanMembran Seloki0% (1)
- Fishew Slide 6-9Dokumen4 halamanFishew Slide 6-9dwi sri ayu annisaBelum ada peringkat
- Li SGD LBM 4Dokumen17 halamanLi SGD LBM 4WJ AjaBelum ada peringkat
- OPTIMIZINGDokumen133 halamanOPTIMIZING미린김Belum ada peringkat
- Struktur Dan Fungsi SelDokumen48 halamanStruktur Dan Fungsi SelBilly Ramadhan100% (1)
- Makalah Transportasi Zat Melalui Membran Sel HewanDokumen16 halamanMakalah Transportasi Zat Melalui Membran Sel HewanSamuel EtoBelum ada peringkat
- SELANGKARAN SELDokumen46 halamanSELANGKARAN SELChoHanaSparkyuBelum ada peringkat
- Struktur Membran SelDokumen12 halamanStruktur Membran SelRobi CioBelum ada peringkat
- BAB I Biologi MolekulerDokumen21 halamanBAB I Biologi MolekulerYane AngelaBelum ada peringkat
- Tugas Struktur SelDokumen8 halamanTugas Struktur Selnznazman_423328299Belum ada peringkat
- Membran Plasma Dan Hubungan Antar SelDokumen23 halamanMembran Plasma Dan Hubungan Antar Selsri yanti100% (3)
- Fungsi Membran PlasmaDokumen3 halamanFungsi Membran PlasmaIrina Natalena Osanti100% (2)
- Cerpen Robohnya Surau KamiDokumen8 halamanCerpen Robohnya Surau Kamizikri arnandaBelum ada peringkat
- Soal Ujian Sekolah Kelas 9 2023 SiswaDokumen11 halamanSoal Ujian Sekolah Kelas 9 2023 SiswaTha Joon Lautner Matsuyama100% (1)
- Biodata ThataDokumen3 halamanBiodata ThataTha Joon Lautner MatsuyamaBelum ada peringkat
- MatrikDokumen1 halamanMatrikTha Joon Lautner MatsuyamaBelum ada peringkat
- Ipi60996 PDFDokumen5 halamanIpi60996 PDFIntanFakhrunNi'amBelum ada peringkat
- 187 317 1 SMDokumen12 halaman187 317 1 SMTha Joon Lautner MatsuyamaBelum ada peringkat
- SURAT PERNYATAAN Publikasi2016v1 - Rev Wadir 1Dokumen1 halamanSURAT PERNYATAAN Publikasi2016v1 - Rev Wadir 1Tha Joon Lautner MatsuyamaBelum ada peringkat
- Kunci Jawaban Uas SD 2019-WPS OfficeDokumen5 halamanKunci Jawaban Uas SD 2019-WPS OfficeTha Joon Lautner MatsuyamaBelum ada peringkat
- Surat Pernyataan Penghasilan FixDokumen1 halamanSurat Pernyataan Penghasilan FixTha Joon Lautner MatsuyamaBelum ada peringkat
- Oret 2Dokumen1 halamanOret 2Tha Joon Lautner MatsuyamaBelum ada peringkat
- Oret 2Dokumen2 halamanOret 2Tha Joon Lautner MatsuyamaBelum ada peringkat
- Oret 2Dokumen2 halamanOret 2Tha Joon Lautner MatsuyamaBelum ada peringkat
- 187 317 1 SM PDFDokumen9 halaman187 317 1 SM PDFTha Joon Lautner MatsuyamaBelum ada peringkat
- AddieDokumen11 halamanAddieTha Joon Lautner MatsuyamaBelum ada peringkat
- Kontrol Genetik Respon ImunDokumen7 halamanKontrol Genetik Respon ImunAnnisa Kecil100% (1)
- Hasil Uji-T CemDokumen1 halamanHasil Uji-T CemTha Joon Lautner MatsuyamaBelum ada peringkat
- UNTUK RESUME KONTROL GENETIKDokumen4 halamanUNTUK RESUME KONTROL GENETIKMuhammad Haidar AmrullahBelum ada peringkat
- CovDokumen13 halamanCovTha Joon Lautner MatsuyamaBelum ada peringkat
- Kontrol Genetik Terhadap Respon Imun BaruDokumen21 halamanKontrol Genetik Terhadap Respon Imun BaruTha Joon Lautner MatsuyamaBelum ada peringkat
- E-Tiket Citilink Anda - No. Pesanan 285048356Dokumen3 halamanE-Tiket Citilink Anda - No. Pesanan 285048356Tha Joon Lautner MatsuyamaBelum ada peringkat
- Enterobacter Asburiae L1 Adalah Bakteri Penginderaan Kuorum Yang Diisolasi Dari Daun SeladaDokumen1 halamanEnterobacter Asburiae L1 Adalah Bakteri Penginderaan Kuorum Yang Diisolasi Dari Daun SeladaTha Joon Lautner MatsuyamaBelum ada peringkat
- Resume 6Dokumen7 halamanResume 6Tha Joon Lautner MatsuyamaBelum ada peringkat
- Refleksi Setelah UAS BiologiDokumen1 halamanRefleksi Setelah UAS BiologiTha Joon Lautner Matsuyama100% (1)
- 2 Baru - En.idDokumen8 halaman2 Baru - En.idTha Joon Lautner MatsuyamaBelum ada peringkat
- Lampiran 1. Angket Validasi ModulDokumen6 halamanLampiran 1. Angket Validasi ModulTha Joon Lautner MatsuyamaBelum ada peringkat
- Bahan Diskusi CHP 17-21Dokumen1 halamanBahan Diskusi CHP 17-21Nur Akbar AbaiBelum ada peringkat
- Pemanfaatan Bakteri Indigen Dalam Bioremidiasi Limbah Cair Puskesmas Di Kota MataramDokumen4 halamanPemanfaatan Bakteri Indigen Dalam Bioremidiasi Limbah Cair Puskesmas Di Kota MataramTha Joon Lautner MatsuyamaBelum ada peringkat
- Refleksi Diri Akhir Semester ThataDokumen4 halamanRefleksi Diri Akhir Semester ThataTha Joon Lautner Matsuyama83% (6)
- Jurnal Belajar 14 - Allvanialista IkalorDokumen6 halamanJurnal Belajar 14 - Allvanialista IkalorTha Joon Lautner MatsuyamaBelum ada peringkat
- Jurnal Belajar 13 - Allvanialista IkalorDokumen7 halamanJurnal Belajar 13 - Allvanialista IkalorTha Joon Lautner MatsuyamaBelum ada peringkat