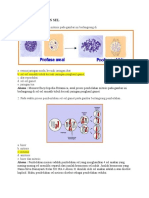
Anda mungkin juga menyukai
- Mikrobiologi Perubatan I: Patogen dan Mikrobiologi ManusiaDari EverandMikrobiologi Perubatan I: Patogen dan Mikrobiologi ManusiaPenilaian: 2.5 dari 5 bintang2.5/5 (2)
- Mikrobiologi Medis I: Patogen dan Mikrobioma ManusiaDari EverandMikrobiologi Medis I: Patogen dan Mikrobioma ManusiaPenilaian: 4 dari 5 bintang4/5 (11)
- JelaskanDokumen4 halamanJelaskanPoetri TeeaBelum ada peringkat
- Studi LiteraturDokumen8 halamanStudi LiteraturNabila MohuneBelum ada peringkat
- Mikrosporogenesis Dan Megasporogenesis 4ADokumen13 halamanMikrosporogenesis Dan Megasporogenesis 4ASonia A100% (1)
- Work Sheet 01Dokumen4 halamanWork Sheet 01Mantabs JiwaBelum ada peringkat
- Makalah Anatomi Kel 6 FixDokumen16 halamanMakalah Anatomi Kel 6 FixSaukiBelum ada peringkat
- Laporan GendasDokumen13 halamanLaporan Gendaskhairun nisak100% (2)
- Pembentukan Dan Perkembangan BungaDokumen7 halamanPembentukan Dan Perkembangan BungaShun NittaBelum ada peringkat
- 049 - Nurul Izzah Fatimah - LKM Siklus Gymnospermae 12Dokumen3 halaman049 - Nurul Izzah Fatimah - LKM Siklus Gymnospermae 12Sayyidah NafisyahBelum ada peringkat
- Laporan EmbtumbDokumen8 halamanLaporan EmbtumbSalmo MooBelum ada peringkat
- Pertanyaan Untuk Kelompok 11Dokumen6 halamanPertanyaan Untuk Kelompok 11kuma edyBelum ada peringkat
- Jelaskan Pengertian Pembuahan Pada TumbuhanDokumen6 halamanJelaskan Pengertian Pembuahan Pada TumbuhanYusuf WahyuBelum ada peringkat
- Spermatogenesis Dan Oogenesis Tugas IndividuDokumen14 halamanSpermatogenesis Dan Oogenesis Tugas Individucindi ahayaBelum ada peringkat
- Tentiran Praktikum Biologi 1 Dan 2Dokumen31 halamanTentiran Praktikum Biologi 1 Dan 2Yucca CameliaBelum ada peringkat
- Pembelahan MeiosisDokumen18 halamanPembelahan MeiosisRulik OktavianiBelum ada peringkat
- Tuton Inisiasi IDokumen18 halamanTuton Inisiasi IfikabonaBelum ada peringkat
- Tugas Per. Hewan-Kelompok 3-1Dokumen32 halamanTugas Per. Hewan-Kelompok 3-1anisa kurniatiBelum ada peringkat
- Bab IDokumen7 halamanBab INofal TahirBelum ada peringkat
- Pengertian Gametogenesis AdalahDokumen4 halamanPengertian Gametogenesis AdalahDiyanti ArfahdBelum ada peringkat
- Kelompok 13Dokumen14 halamanKelompok 13Asnawar RustamBelum ada peringkat
- TUGAS INDIVIDU - Benih. DHIDYDokumen4 halamanTUGAS INDIVIDU - Benih. DHIDYWhey DedewBelum ada peringkat
- Life Cycle of Pine Dan ManfaatnyaDokumen7 halamanLife Cycle of Pine Dan ManfaatnyaAulia MasyitohBelum ada peringkat
- Embrio AmphioxusDokumen15 halamanEmbrio AmphioxusrhyphamBelum ada peringkat
- Latihan Soal Pembelahan Sel 2021-2022Dokumen7 halamanLatihan Soal Pembelahan Sel 2021-2022Chandra PutraBelum ada peringkat
- Megasporogenesis Dan Embriogenesis PinusDokumen4 halamanMegasporogenesis Dan Embriogenesis PinusLaila Alvi NurinBelum ada peringkat
- Analisis MiosisDokumen17 halamanAnalisis Miosiskhairulnasution678100% (1)
- AngiospermaeDokumen4 halamanAngiospermaeRunoffBelum ada peringkat
- Mikrosporogenesis Dan MakrosporogenesisDokumen6 halamanMikrosporogenesis Dan MakrosporogenesisAyu Rizki IkhsaniBelum ada peringkat
- FERTILISASIDokumen23 halamanFERTILISASINisa Sifa SabilaBelum ada peringkat
- Tugas 1Dokumen11 halamanTugas 1Neni YuningsihBelum ada peringkat
- Putik Bunga Lili Memiliki Bagian Ovul Sebagai Tempat Pembentukan Gametofit BetinaDokumen2 halamanPutik Bunga Lili Memiliki Bagian Ovul Sebagai Tempat Pembentukan Gametofit BetinaSuparjo RustamBelum ada peringkat
- Developmental Biology 7th Ed SF Gilbert 228 263Dokumen36 halamanDevelopmental Biology 7th Ed SF Gilbert 228 263edy prasetyoBelum ada peringkat
- TUGAS IDK ReproduksiDokumen16 halamanTUGAS IDK ReproduksiTania PangestuBelum ada peringkat
- Angiospermae UmumDokumen12 halamanAngiospermae UmumPonti PontiBelum ada peringkat
- Tugas 2Dokumen3 halamanTugas 2VaniBelum ada peringkat
- Gametogenesis Pada Hewan Dan TumbuhanDokumen4 halamanGametogenesis Pada Hewan Dan TumbuhanFitria IdaBelum ada peringkat
- TaizDokumen14 halamanTaizmediaku123Belum ada peringkat
- CycasDokumen26 halamanCycasLeni MileniBelum ada peringkat
- Pemasakan Organ Kelamin BungaDokumen10 halamanPemasakan Organ Kelamin BungaEmma Femi P.Belum ada peringkat
- Metagenesis Tumbuhan LumutDokumen13 halamanMetagenesis Tumbuhan LumutMuzdar HmcBelum ada peringkat
- Biolog IDokumen6 halamanBiolog IAndreas MalvinBelum ada peringkat
- Jonathan Ecclecia Polandos-1Dokumen5 halamanJonathan Ecclecia Polandos-1jonathan polandosBelum ada peringkat
- Fertilisasi MakalahDokumen34 halamanFertilisasi MakalahNining Nurnaningsih33% (3)
- Sporagenesis Dan GametogenesisDokumen17 halamanSporagenesis Dan GametogenesisArlinda Puspita SariBelum ada peringkat
- Dokumen - Tips Acara PraktikumDokumen4 halamanDokumen - Tips Acara Praktikumtika handayaniBelum ada peringkat
- Mikrosporogenesis 130921060839 Phpapp02Dokumen3 halamanMikrosporogenesis 130921060839 Phpapp02rinaldy husainBelum ada peringkat
- Tugas Teknologi BenihDokumen11 halamanTugas Teknologi BenihDian SusantiBelum ada peringkat
- Mitosis Dan MeiosisDokumen4 halamanMitosis Dan MeiosisSheilly Olivia MarcelindaBelum ada peringkat
- Siti Aminah - 22334754 - Tugas 5 Biologi SelDokumen5 halamanSiti Aminah - 22334754 - Tugas 5 Biologi Selpkl cipinangcempedakBelum ada peringkat
- TUGAS 5 BioselDokumen5 halamanTUGAS 5 Bioselpkl cipinangcempedakBelum ada peringkat
- Polen MeiosisDokumen5 halamanPolen MeiosisKsatria BimaBelum ada peringkat
- Wildan Sahari - PRAK 6Dokumen11 halamanWildan Sahari - PRAK 6wildan sahariBelum ada peringkat
- Makalah Perkembangan Embrio Mamalia AriadnaDokumen9 halamanMakalah Perkembangan Embrio Mamalia Ariadnaariadna safitri100% (2)
- Mitosis Dan MeiosisDokumen4 halamanMitosis Dan MeiosisMirza Faris RahmanBelum ada peringkat
- Mitosis Dan MeiosisDokumen4 halamanMitosis Dan MeiosisYoZa JunNovBelum ada peringkat
- 2 Membaca Dan Merangkum TulisanDokumen40 halaman2 Membaca Dan Merangkum TulisanRahma KartikaBelum ada peringkat
- 4637 11394 1 SMDokumen9 halaman4637 11394 1 SMRahma KartikaBelum ada peringkat
- Rahma Eka Kartika - K4320066 - A - Revisi BannerDokumen1 halamanRahma Eka Kartika - K4320066 - A - Revisi BannerRahma KartikaBelum ada peringkat
- RPP Struktur Dan Fungsi Sel Pada SistemDokumen9 halamanRPP Struktur Dan Fungsi Sel Pada SistemHairul RachmanBelum ada peringkat
- Mekanisme & Kisi-Kisi Materi Uas 2023Dokumen3 halamanMekanisme & Kisi-Kisi Materi Uas 2023Rahma KartikaBelum ada peringkat
- Aulia Nugrahani Sutrisna - K4320011 - MakroalgaDokumen40 halamanAulia Nugrahani Sutrisna - K4320011 - MakroalgaRahma KartikaBelum ada peringkat
- Materi EmwanDokumen180 halamanMateri EmwanRahma KartikaBelum ada peringkat
- Rahma Eka Kartika - K4320066 - C - RPP Sistem Pencernaan (Role Playing)Dokumen15 halamanRahma Eka Kartika - K4320066 - C - RPP Sistem Pencernaan (Role Playing)Rahma KartikaBelum ada peringkat
- Kelompok 5A - Uji Rata-Rata Data Berpasangan Dan Tidak BerpasanganDokumen21 halamanKelompok 5A - Uji Rata-Rata Data Berpasangan Dan Tidak BerpasanganRahma KartikaBelum ada peringkat
- Perhitungan Jumlah BakteriDokumen11 halamanPerhitungan Jumlah BakteriMarifatul Fajriyah100% (1)
- Buku Album Makrab 2020Dokumen3 halamanBuku Album Makrab 2020Rahma KartikaBelum ada peringkat
- Laporan Praktikum Perhitungan Jumlah MikrobaDokumen15 halamanLaporan Praktikum Perhitungan Jumlah MikrobaRahma KartikaBelum ada peringkat
- Sistem Saraf - Kelompok 3Dokumen36 halamanSistem Saraf - Kelompok 3Rahma KartikaBelum ada peringkat
- Memperkokoh Peran Mahasiswa Dalam Menangkal Radikalisme Dan Terorisme Melalui Penerapan PancasilaDokumen15 halamanMemperkokoh Peran Mahasiswa Dalam Menangkal Radikalisme Dan Terorisme Melalui Penerapan PancasilaRahma KartikaBelum ada peringkat
- MountingDokumen1 halamanMountingRahma KartikaBelum ada peringkat
- Rahma Eka Kartika - K4320066 - Laporan Praktikum Mikrobiologi 4Dokumen39 halamanRahma Eka Kartika - K4320066 - Laporan Praktikum Mikrobiologi 4Rahma KartikaBelum ada peringkat
- Permasalahan Ekosistem BakauDokumen4 halamanPermasalahan Ekosistem BakauRahma KartikaBelum ada peringkat
- DokumenDokumen1 halamanDokumenRahma KartikaBelum ada peringkat
- Uji Data Tidak Normal Mann WhitneyDokumen2 halamanUji Data Tidak Normal Mann WhitneyRahma KartikaBelum ada peringkat
- Webinar PancasilaDokumen11 halamanWebinar PancasilaRahma KartikaBelum ada peringkat
- Aulia Nugrahani Sutrisna - K4320011 - A - Laprak AHH 1Dokumen26 halamanAulia Nugrahani Sutrisna - K4320011 - A - Laprak AHH 1Rahma KartikaBelum ada peringkat
- Rahma Eka Kartika - K4320066 - A - Tugas Biostat Pertemuan 11Dokumen11 halamanRahma Eka Kartika - K4320066 - A - Tugas Biostat Pertemuan 11Rahma KartikaBelum ada peringkat
- Alat Dan Bahan TaksidermiDokumen1 halamanAlat Dan Bahan TaksidermiRahma KartikaBelum ada peringkat
- Riannisa Shafira - K4320069 - A - Laporan Resmi AHH - Jaringan DasarDokumen22 halamanRiannisa Shafira - K4320069 - A - Laporan Resmi AHH - Jaringan DasarRahma KartikaBelum ada peringkat
- Permasalahan Ekosistem BakauDokumen4 halamanPermasalahan Ekosistem BakauRahma KartikaBelum ada peringkat
- Kelompok Tax Reptil - Kemikal Protokol ProsedurDokumen9 halamanKelompok Tax Reptil - Kemikal Protokol ProsedurRahma KartikaBelum ada peringkat
- Kelompok 1 Prak AHHDokumen14 halamanKelompok 1 Prak AHHRahma KartikaBelum ada peringkat
- Media AudioDokumen13 halamanMedia AudioRahma KartikaBelum ada peringkat
- Fix Rangkuman Tugas Telkum 11Dokumen2 halamanFix Rangkuman Tugas Telkum 11Rahma KartikaBelum ada peringkat