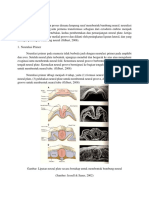
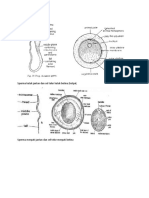
Anda mungkin juga menyukai
- Penghapusan Urutan Intron Dengan Penyambungan RNADokumen4 halamanPenghapusan Urutan Intron Dengan Penyambungan RNAGufron AlifiBelum ada peringkat
- Modifikasi Pasca TranskripsiDokumen11 halamanModifikasi Pasca TranskripsiBinazir Tuza Qiyah Ma'rufahBelum ada peringkat
- TRANSLASIDokumen8 halamanTRANSLASIAmanda Icush Febriyani0% (1)
- Resume 7 TranslasiDokumen5 halamanResume 7 TranslasiGesty MBelum ada peringkat
- Resume Plasmid Dan EpisomeDokumen5 halamanResume Plasmid Dan EpisomeMickz KresnhaBelum ada peringkat
- Resume Mutasi Kromosom Bab 5 Dan Bab 6Dokumen10 halamanResume Mutasi Kromosom Bab 5 Dan Bab 6Lyana BerlyBelum ada peringkat
- Ekspresi Gen Pada Eukariot GenetikaDokumen22 halamanEkspresi Gen Pada Eukariot GenetikaLailyrahma100% (3)
- Transposable Element GeneticDokumen23 halamanTransposable Element GeneticOki Osaka100% (1)
- Makalah Teori Evolusi DarwinDokumen10 halamanMakalah Teori Evolusi DarwinSyadiq AbbasBelum ada peringkat
- Mutasi Bab 4 5 N 6Dokumen6 halamanMutasi Bab 4 5 N 6Muhammad Shölih Qoyyim0% (1)
- Tugas 1Dokumen4 halamanTugas 1intan yunanda100% (1)
- REGULASI GEN EUKARIOTDokumen9 halamanREGULASI GEN EUKARIOTDewi AnisaBelum ada peringkat
- Rqa Genetika 2Dokumen5 halamanRqa Genetika 2Permata Windra DeasmaraBelum ada peringkat
- BIOINDUKATOR AIRDokumen10 halamanBIOINDUKATOR AIRReza AnisaBelum ada peringkat
- Resume EkstrakromosomalDokumen9 halamanResume EkstrakromosomalAstrinoPurmannaBelum ada peringkat
- Plasmid Dan EpisomDokumen6 halamanPlasmid Dan EpisomDevintaBelum ada peringkat
- Rqa 3Dokumen5 halamanRqa 3pi nitBelum ada peringkat
- Regulasi Ekspresi Gen Pada Makhluk Hidup ProkariotikDokumen5 halamanRegulasi Ekspresi Gen Pada Makhluk Hidup ProkariotikSITI WIDYAWATIBelum ada peringkat
- Transkripsi ProkariotikDokumen5 halamanTranskripsi ProkariotikRiris Novia AzemiBelum ada peringkat
- Makalah Ekologi (Faktor Biotik)Dokumen14 halamanMakalah Ekologi (Faktor Biotik)Ghio NajoanBelum ada peringkat
- TRANSPOSABLE ELEMENTS GENETICSDokumen2 halamanTRANSPOSABLE ELEMENTS GENETICShilhilloBelum ada peringkat
- Laju Mutasi Dan Deteksi MutasiDokumen3 halamanLaju Mutasi Dan Deteksi MutasiBinazir Tuza Qiyah Ma'rufahBelum ada peringkat
- Plasmid dan EpisomDokumen5 halamanPlasmid dan EpisomNovan AdhiBelum ada peringkat
- MUTASI BAGIAN 2Dokumen17 halamanMUTASI BAGIAN 2Nurul YanuarsihBelum ada peringkat
- Plasmid Dan EpisomeDokumen12 halamanPlasmid Dan EpisomeDasrianiBelum ada peringkat
- Implantasi & Plasentasi FixDokumen38 halamanImplantasi & Plasentasi FixAnisa RositaBelum ada peringkat
- Regulasi Ekspresi Gen EukariotDokumen16 halamanRegulasi Ekspresi Gen EukariotNur Raiyan JannahBelum ada peringkat
- Kel 10 - Resume3 - Replikasi DNADokumen18 halamanKel 10 - Resume3 - Replikasi DNAThania AyuBelum ada peringkat
- Resume Translasi Dan Kode GenetikDokumen7 halamanResume Translasi Dan Kode GenetikIfa AuliaBelum ada peringkat
- TRANSLASIDokumen7 halamanTRANSLASIFaiza ImawatiBelum ada peringkat
- Modifikasi Pasca TranskripsiDokumen7 halamanModifikasi Pasca TranskripsiMita Larasati100% (1)
- Resume Mutasi BaruDokumen8 halamanResume Mutasi BaruhosniyahBelum ada peringkat
- Pembentukan Pankreas, Trakea Dan Paru-Paru, Dan GinjalDokumen2 halamanPembentukan Pankreas, Trakea Dan Paru-Paru, Dan GinjalRIFDA AHADINABelum ada peringkat
- Translasi Dan Kode GenetikDokumen8 halamanTranslasi Dan Kode GenetikFiqih Dewi MaharaniBelum ada peringkat
- KECENDERUNGAN KAWIN DROSOPHILADokumen38 halamanKECENDERUNGAN KAWIN DROSOPHILADiah Ajeng MustikariniBelum ada peringkat
- Makalah Subkelas ThaliceaeDokumen11 halamanMakalah Subkelas ThaliceaeRizki Inggil IIBelum ada peringkat
- SITOSKELETON DAN HUBUNGAN ANTAR SELDokumen53 halamanSITOSKELETON DAN HUBUNGAN ANTAR SELNUR ASMA RIZKYBelum ada peringkat
- REGENERASI IKANDokumen3 halamanREGENERASI IKANIfa AuliaBelum ada peringkat
- Evolusi BurungDokumen6 halamanEvolusi BurungFichar Fitrawan SyarbinBelum ada peringkat
- MUTASI GENETIKADokumen11 halamanMUTASI GENETIKAKarina Nur'ainiBelum ada peringkat
- TranslasiDokumen12 halamanTranslasiElsa NoviaBelum ada peringkat
- AnnelidaDokumen3 halamanAnnelidaVarian ForturozyBelum ada peringkat
- Metamorfosis KatakDokumen15 halamanMetamorfosis KatakUmmu HalimahBelum ada peringkat
- Rekayasa GenetikaDokumen8 halamanRekayasa GenetikaDewa Ayu SwaratriBelum ada peringkat
- Rolling Circle ReplicationnDokumen11 halamanRolling Circle ReplicationnmamonBelum ada peringkat
- TranslasiDokumen9 halamanTranslasiSinta HandayaniBelum ada peringkat
- Laporan Praktikum 6 Metode KuadratDokumen15 halamanLaporan Praktikum 6 Metode KuadratFirda Ayu WahyuniBelum ada peringkat
- Proses Intesis ProteinDokumen16 halamanProses Intesis ProteinNisahBelum ada peringkat
- GENETIKA KELAMINDokumen24 halamanGENETIKA KELAMINGizella Ayu WilantikaBelum ada peringkat
- LKS SISTEM SARAF 1Dokumen8 halamanLKS SISTEM SARAF 1Koko MurdiantoBelum ada peringkat
- RESUME Macam MutasiDokumen3 halamanRESUME Macam MutasiyoanandaBelum ada peringkat
- Interaksi Gen dan Ekspresi SifatDokumen9 halamanInteraksi Gen dan Ekspresi SifatAdauchi EndiBelum ada peringkat
- Regulasi Ekspresi Gen Pada EukariotikDokumen12 halamanRegulasi Ekspresi Gen Pada EukariotikChad SharpBelum ada peringkat
- Makalah KomunikasiDokumen15 halamanMakalah KomunikasiAri ZusriadiBelum ada peringkat
- Siklus Glioksilat Dan GlukoneogenesisDokumen2 halamanSiklus Glioksilat Dan GlukoneogenesisDhea DesitaBelum ada peringkat
- Struktur Materi Genetik EditDokumen11 halamanStruktur Materi Genetik EditAkim Abdi MarsBelum ada peringkat
- KELAS AMPHIBIADokumen10 halamanKELAS AMPHIBIAhildaBelum ada peringkat
- Elongasi Rantai Rna Dan PenambahanDokumen3 halamanElongasi Rantai Rna Dan PenambahanBundaana CLuuk BaaBelum ada peringkat
- Modifikasi Pasca TranskripsiDokumen4 halamanModifikasi Pasca Transkripsiismiatul0% (1)
- Modifikasi Pasca TranskripsiDokumen7 halamanModifikasi Pasca TranskripsiAbdulfattah NoorBelum ada peringkat
- DETEKSI METABOLIT SEKUNDERDokumen1 halamanDETEKSI METABOLIT SEKUNDERida nurpitaBelum ada peringkat
- Halaman I II III IV, Tinggal Ngasih HalamanDokumen3 halamanHalaman I II III IV, Tinggal Ngasih Halamanida nurpitaBelum ada peringkat
- Resume Istilah ParasitologiDokumen20 halamanResume Istilah Parasitologiida nurpitaBelum ada peringkat
- Rujukan KH AvesDokumen8 halamanRujukan KH Avesida nurpitaBelum ada peringkat
- Diversitas Antibod1 1Dokumen3 halamanDiversitas Antibod1 1ida nurpitaBelum ada peringkat
- Pembahasan PHDokumen7 halamanPembahasan PHida nurpitaBelum ada peringkat
- Kata PengantarDokumen2 halamanKata Pengantarida nurpitaBelum ada peringkat
- Skrining FitokimiaDokumen1 halamanSkrining Fitokimiaida nurpitaBelum ada peringkat
- PudjiadiDokumen1 halamanPudjiadiida nurpitaBelum ada peringkat
- Tri TerpenoidDokumen5 halamanTri Terpenoidida nurpitaBelum ada peringkat
- Factsheet WWF Indonesia Rangkong Badak PDFDokumen2 halamanFactsheet WWF Indonesia Rangkong Badak PDFida nurpitaBelum ada peringkat
- 4Dokumen4 halaman4ida nurpitaBelum ada peringkat
- Ekinodermata Kelas dan Ciri MorfologiDokumen15 halamanEkinodermata Kelas dan Ciri Morfologiida nurpita0% (1)
- DETEKSI METABOLIT SEKUNDERDokumen1 halamanDETEKSI METABOLIT SEKUNDERida nurpitaBelum ada peringkat
- Kelompok Ikan - Kelompok 1Dokumen58 halamanKelompok Ikan - Kelompok 1ida nurpitaBelum ada peringkat
- Resume Materi Griffith DKKDokumen3 halamanResume Materi Griffith DKKida nurpitaBelum ada peringkat
- Daftar Rujukan Tekanan Jantung Dan VenaDokumen5 halamanDaftar Rujukan Tekanan Jantung Dan Venaida nurpitaBelum ada peringkat
- Poin 2Dokumen5 halamanPoin 2ida nurpitaBelum ada peringkat
- HolothuroideaDokumen2 halamanHolothuroideaida nurpitaBelum ada peringkat
- Peranan MikroDokumen4 halamanPeranan Mikroida nurpitaBelum ada peringkat
- Pembahasan Gerak BakteriDokumen2 halamanPembahasan Gerak Bakteriida nurpitaBelum ada peringkat
- Daftar PustakaDokumen3 halamanDaftar Pustakaida nurpitaBelum ada peringkat
- Laporan Pith Fall Kelompok 23Dokumen8 halamanLaporan Pith Fall Kelompok 23ida nurpitaBelum ada peringkat
- NEURULASI PRIMER DAN SEKUNDERDokumen5 halamanNEURULASI PRIMER DAN SEKUNDERida nurpitaBelum ada peringkat
- Sperma Katak Jantan Dan Sel Telur Katak BetinaDokumen2 halamanSperma Katak Jantan Dan Sel Telur Katak Betinaida nurpitaBelum ada peringkat
- Diskusi, Kesipulan, Dapus ALTDokumen1 halamanDiskusi, Kesipulan, Dapus ALTida nurpitaBelum ada peringkat
- KLASIFIKASI IKAN BERTULANG KERASDokumen16 halamanKLASIFIKASI IKAN BERTULANG KERASida nurpitaBelum ada peringkat
- Operonlakpdf PDFDokumen5 halamanOperonlakpdf PDFida nurpitaBelum ada peringkat
- Asteroidea OphiuroideaDokumen7 halamanAsteroidea Ophiuroideaida nurpitaBelum ada peringkat
- Peranan MikroDokumen4 halamanPeranan Mikroida nurpitaBelum ada peringkat